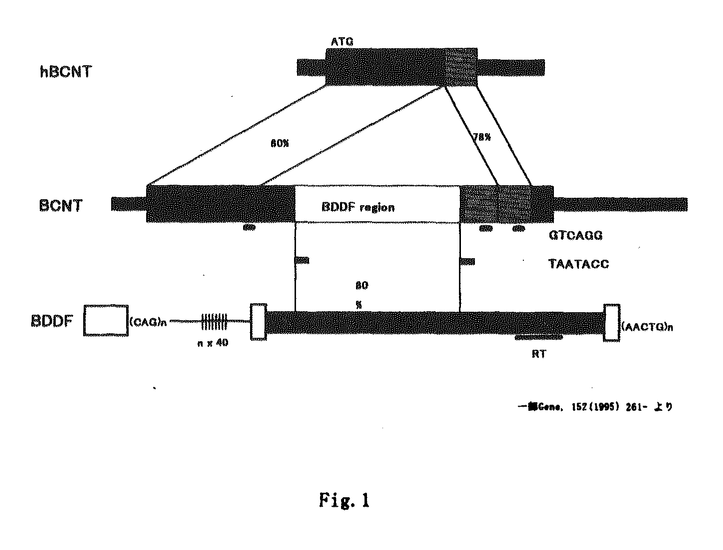
本論文は,種々の哺乳類で保存され,進化論上非常に興味深い構造を有した新規蛋白質の発見及びその生化学的特徴を報告したものである。解析は主に分子生物学的手法を用いた遺伝子の同定と,蛋白質自身の性質に関して行われた。以下にそのようやくを示す。 真核生物のゲノム中には反復配列と呼ばれる配列が、ゲノムあたり105-106コピー存在し、全塩基数の数パーセントを占めている。哺乳類の反復配列は、300bp程度の長さで、蛋白質をコードしていないSINE(Short INterspersed Element)と、数kbpの長さを持ち逆転写酵素様の蛋白質をコードしているLINE(Long INterspersed Element)の2種類に分類することができる。霊長類のAlu配列に代表される前者は、その起源がtRNAや7SL RNAといった内在性のRNA分子であると考えられている。後者は、そのコンセンサス配列に2個のORF(ORF1,ORF2)を持ち、自身のORF2にコードされる逆転写酵素によって増幅するものと考えられている。 これらの配列のごく一部のコピーは実際にORFを持っており、蛋白になることが示されたものもある。しかしながら、ほとんどのコピーは、元々ORFをコードしていないか、変異が起こった結果、転写、または翻訳が正常に起こらないために蛋白質として発現していない。従ってこれらの配列の増幅する元になるコピーは、非常に限られたものであるか、あるいはただ1つ(Master Gene)であるかもしれない。 このような点が明らかになった段階であって、ゲノム中の大きな部分を占める反復配列の機能はいまだに分かっていない。従って、その存在意義を示すことはゲノムの構造、進化を研究する上で非常に意義深い。 低分子量G蛋白質の代表であり原癌遺伝子であるrasを負に制御する因子であるrasGAP(rasGTPase Activating Protein)ファミリーの一員である に対するモノクローナル抗体が、ウシ脳の抽出物中の他の因子と反応することを見いだした。この蛋白質はSDS-PAGE上の見かけの大きさが97kDa(p97)であることなど、と良く似た性質を持っていた。しかし、このモノクローナル抗体がとは弱くしか反応しないため、p97は新たなrasGAPファミリーの一員であると予想し,このcDNAをクローニングすることとした。 に対するモノクローナル抗体が、ウシ脳の抽出物中の他の因子と反応することを見いだした。この蛋白質はSDS-PAGE上の見かけの大きさが97kDa(p97)であることなど、と良く似た性質を持っていた。しかし、このモノクローナル抗体がとは弱くしか反応しないため、p97は新たなrasGAPファミリーの一員であると予想し,このcDNAをクローニングすることとした。 最初に,ウシcDNAの発現ライブラリーを,この抗体でスクリーニングすることによりクローニングすることを試みたが、ヒトのLINEであるL1に相同性の高いクローンが得られ、p97に相当すると思われるcDNAはクローニングできなかった。 そこで,このモノクローナル抗体を用いてp97蛋白質を精製し,その部分アミノ酸配列を決定し,これを基にPCR法によりそのcDNAの一部をクローニングし,RACE法によりcDNAの両端をクローニングした。 この遺伝子は予想に反しrasGAPファミリーの一員ではなかったが、新規な蛋白質をコードしていた。遺伝子の配列を解析した結果、592残基のアミノ酸配列をコードしており、そのORF中央部の約800塩基対は、ウシで発見された反復配列であるBDDF(Bovine Dimer Driven Family)と80%の相同性を示すことが分かった。 そこで、このような配列を持つ蛋白質がcDNAとしてクローニングされたことがPCR等の誤りであるかどうかを考察した。まず、プローブの合成の目的のみではなく、数多くのペプチドのアミノ酸配列を決定したところ、実際に決定されたアミノ酸配列の中に反復配列部位に相当するものが存在することから、決定されたcDNA配列が正しい遺伝子より由来するものだと確信された。さらに、PCRを使わずに,反復配列領域を含んだcDNAがクローニングされるかどうかを調べるため,ファージライブラリーを,反復配列領域を含まないプローブでスクリーニングした。その結果得られたクローンは全て前記の結果と矛盾せず、反復配列領域を含んでいた。 さらに、データベースを検索した結果,ヒトやマウスのcDNAプロジェクトの中に、このcDNAと非常に高い相同性をもつものが存在することを検出した。そこで,ウシの場合と同様に,2通りの方法でヒトのcDNAをクローニングした。即ちESTプロジェクトよりフラグメントを入手しその塩基配列を再決定しこれを基にRACE法により全長をクローニングした。またウシのcDNA配列の内反復配列領域を含まないN端側の領域をプローブにヒトcDNAのファージライブラリーをスクリーニングした。得られたcDNAはデータベースより得られた配列および上記でRACEにより得られた配列とほぼ一致した。得られたcDNAはウシの遺伝子とORFに関して、80%の相同性を持っていたが、反復配列相同領域およびこれに続く40アミノ酸残基の領域を欠失していた(Fig.1)。そこで、この新規蛋白質を明らかにした経過をもとに,半牛半人の神Bucentaurから、BCNTと命名した。また、ヒトのホモログをhBCNTと呼ぶこととした。 これらの結果より推察されることは,BCNTは少なくとも哺乳類では保存されており何らかの重要な機能を果たしているが、ウシでは,反復配列の挿入が起こった結果,蛋白質産物のサイズが大きくなっている。ただし、種間の保存性の高さ、挿入が起きても、それ以降のフレームがずれることなくORFが続いていることなどから、その機能が損なわれていないことが予想された。 そこで、ウシで反復配列領域のないヒト型のBCNTの有無を調べるため以下の実験を行った。まず、ジェノミックサザンを行ったところ、BCNTは恐らくゲノム中に単一コピーとして存在していることが示された。 また、p97蛋白質の精製および同定に用いた抗ウシBCNT抗体のエピトープを決定した。その結果,用いた全てのモノクローナル抗体は同じBCNTの反復配列領域よりもN端側の部位を認識していることが判明した。この抗体を用いた免疫ブロッティングの結果,97kDaより小さな分子は認められなかった。これらの事実より,ウシでは遺伝子および蛋白質レベルで反復配列領域の欠如した分子型のBCNTが存在する可能性が極めて低いことが明らかとなった。 以上の結果、BCNTには機能の欠損を起こさずに反復配列の挿入が起こったことが強く示唆された。このように,大きな構造変化が,蛋白質の機能を欠損することなく起こったことが示されるのは非常に珍しい例であり、進化論的に非常に興味深い研究対象といえる。 BCNTは、N端に,酸性アミノ酸残基に富みカゼインカイネスIIによる燐酸化のコンセンサスを持つ配列,C端に、40アミノ酸残基からなる繰り返し配列(IR;Intramolecular Repeat)を持つ,興味深い構造を持っており,その機能を明らかにすることは,進化論的考察を含め,非常に学問的意義が大きい。そのため、BCNTの遺伝子の同定とそれに続く諸性質の解析は重要な学術的な貢献をするものと認められる。このため、信國宇洋は博士(理学)の学位を受けるのに十分な資格があるものと判断された。 |