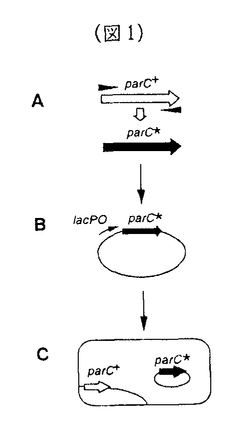
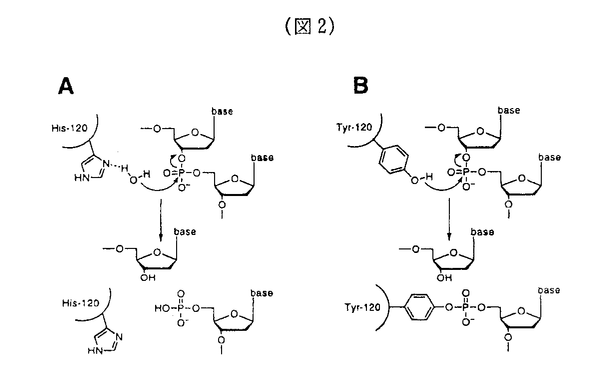
大腸菌DNAトポイソメラーゼIVは、染色体の分配において重要な役割を持つ酵素であり、環状DNAの複製の際に生じるDNA間の絡まりをほぐす働きをしている。II型トポイソメラーゼの反応機構の複雑さのために、トポイソメラーゼの反応機構の詳細なメカニズムは未だに明かではない。本論文は、DNAトポイソメラーゼIVに関する遺伝学的な解析によって反応機構の解明に迫ることを目的としたものである。 本論文の実験の結果及び考察は、第2章にまとめられている。第2章第1節では、トポイソメラーゼIVの反応過程の様々なステップに欠損を持つ変異株の単離を目的として、その優性致死突然変異株を単離する方法を開発したことと、得られた変異株の一般的な性質をしらべ、目的とした突然変異株が得られたことを確認した。複雑な反応過程を持つ酵素について優性致死突然変異株を単離するこの方法は、他に類を見ない新しいものである。 第2章第2節では、優性致死突然変異株parC3について、SOS応答の誘導を測定するアッセイ系を確立し、parC3変異の導入によって細胞内でDNA切断が起きることを示した。この結果から、変異型トポイソメラーゼIVがDNA鎖の再結合に欠損を持つことがわかった。また、parC3遺伝子の変異部位を同定し、119番目のアルギニンがシステインに変わっていることを示した。 第2章第3節では、トポイソメラーゼIVのATPase活性ドメインを持つParEサブユニットに関する優性致死突然変異株を単離し、変異によってSOS応答が誘導されることを示した。さらに、ATP結合ドメインに対して部位特異的に変異を導入した変異株を単離し、やはり細胞内でDNA切断を引き起こすことをin vivoで示した。 第2章第3節では、優性致死突然変異株parC10を単離し、この変異株がin vivoでSOS応答を誘導していることを示した。さらに、その変異は、トポイソメラーゼIVがDNAと共有結合をつくるための活性中心とされる120番目のチロシン残基がヒスチジン残基に置換されていることを明らかにした。さらに、変異型ParC10蛋白質を精製して、in vitroで変異型トポイソメラーゼIVを再構成し、酵素活性を測定した。その結果、この変異型トポイソメラーゼは、野生型酵素とは異なって、ATP非依存的にDNAの一本鎖切断を起こすことと、その反応産物がトポイソメラーゼと共有結合していないことを明らかにした。このことから、変異型トポイソメラーゼIVが、DNA鎖切断のステップに欠損を持っていると推定し、ヒスチジン残基に水素結合した水分子がDNA鎖を求核攻撃するというモデルで、変異型トポイソメラーゼIVの活性を説明した。 以上の結果は、野生型ParC蛋白質においては、120番目のチロシン残基がDNAの切断の過程に重要な役割を持つことを遺伝学的に証明したものである。同時に、チロシン残基のフェノール性水酸基の求核性を高める機構がParC蛋白質の活性部位近傍に存在する、ということが示唆された。さらに、119番目のアルギニン残基は、切断の再結合の過程に重要な役割を持つことを示した。 本論文は、II型トポイソメラーゼの反応機構の各ステップを遺伝学的に解析する方法の端緒を開いたものであり、この酵素の高次構造の解明と相まって、酵素の構造と機能の問題の解明の重要なステップとなるものであり、この領域へ大きく寄与したものと考えられる。よって、博士(理学)の学位を授与するに値することを認める。 なお、本論文は、加藤潤一博士、池田日出男博士との共同研究であるが、論文提出者が主体となって分析及び検証を行ったもので、論文提出者の寄与が十分であると判断する。 |