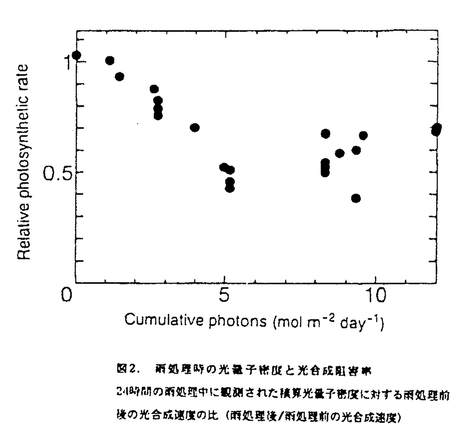
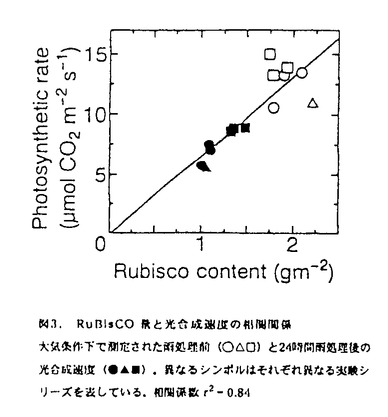
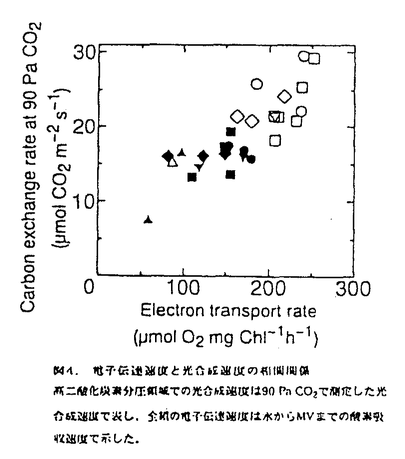
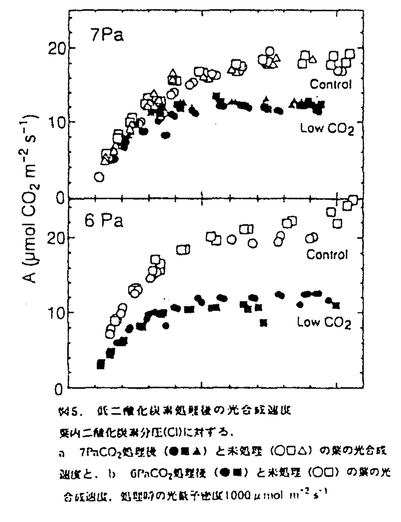
本論文は三章からなり、第一章には長期間の濡れが葉の光合成や気孔反応にどのような影響を与えるか、第二章には葉の濡れによって引き起こされた光合成の阻害のメカニズムについて、そして第三章には二酸化炭素欠乏の影響について述べられている。 第一章では、葉の濡れに対する気孔開閉反応とともに分という時間スケールで光合成が変化することを明らかにするとともに、光合成の阻害がどれくらいの時間、濡れに晒されると起こるのか、そして処理時の光環境の阻害への影響など、濡れによる光合成阻害の条件を特定している。濡れに対する光合成の反応が気孔反応と同じように変化し、その一時的な濡れによる光合成の低下が可逆的なのに対して、6時間以上の葉の濡れによって引き起こされる光合成の阻害は不可逆的であり、光合成研究の長い歴史の中で見逃されてきた全く新しい特性の発見である。そして、その阻害が処理時の積算光量がある程度以上にならないと誘発されないことも明らかにしている。 第二章では、濡れによる光合成阻害のメカニズムについて、ふたつの要因に分けて議論している。ひとつは炭酸固定酵素ルビスコの分解で、酵素の初期活性、最大活性とともに、タンパク量やウェスタンブロットでの確認など適切な操作を行って、リブロース二リン酸カルボキシラーゼ(RuBisCo)が24時間の濡れによって処理前の約半分にまで減少し得ることを立証している。これは、脅威的な速さであり、その分解の詳細なメカニズムをここでは明らかにしていないとはいえ、濡れによってこのようなタンパク質の分解が起こることはひとつの発見であると高く評価できる。もうひとつの、RuBisCoの減少に前もって起こる光合成阻害の原因を電子伝達速度の低下と特定している。これについては、クロロフィル蛍光の経時変化や電子伝達速度の計測によって、これまで知られていたような光化学系Iや系IIの光傷害ではなく、全体としての電子伝達速度が低下していることなどから系Iと系IIの間の部位の傷害であることを推察している。間接的な結果からチトクロームb/f複合体の傷害ではないかと予測しているが、その可能性については今後の検証が必要であろう。しかし、これは全く新しいタイプの光阻害であり、阻害部位の特定は光エネルギーの需要供給のバランスの制御メカニズムを考える上でも大変興味深い。 第三章では、濡れの直接の原因として二酸化炭素欠乏の可能性を考え、濡れた葉に予想される二酸化炭素分圧で植物体を処理し、その光合成や酵素活性、クロロフィルの蛍光など、第二章で検証した濡れによる阻害部位を比較している。光合成速度は二酸化炭素分圧6Paの処理によってどの測定二酸化炭素分圧でも光合成速度が低下し、濡れによる阻害を受けた葉と同様に光化学系Iでも系IIにも傷害がない状態で電子伝達活性が低下していた。しかし、濡れによる阻害ではRuBisCoの分解があったが二酸化炭素欠乏処理ではRuBisCoの活性低下にとどまり、分解は見られなかった。このことは、濡れによる阻害の直接的な原因が二酸化炭素欠乏だけでは説明されないことを示している。二酸化炭素欠乏によって低下したRuBisCoの活性化率低下が分解につながるのかどうかという問題は、その分野でも意見の分かれる難しい問題で本論文の範囲を超えているが、全般的に、しっかりとした目的意識のもとに実験を組み立てる独創性と、得られた結果からの考察の緻密さは群を抜いていると評価できる。 なお、本論文第一章は寺島一郎氏との、第二章の前半は寺島一郎氏と臼田秀明氏との、また後半と第三章は園池公毅氏と渡邊昭との共同研究であるが、論文提出者が主体となって分析及び検証を行ったもので、論文提出者の寄与が十分であると判断する。 よって、博士(理学)の学位を授与できると認める。 | 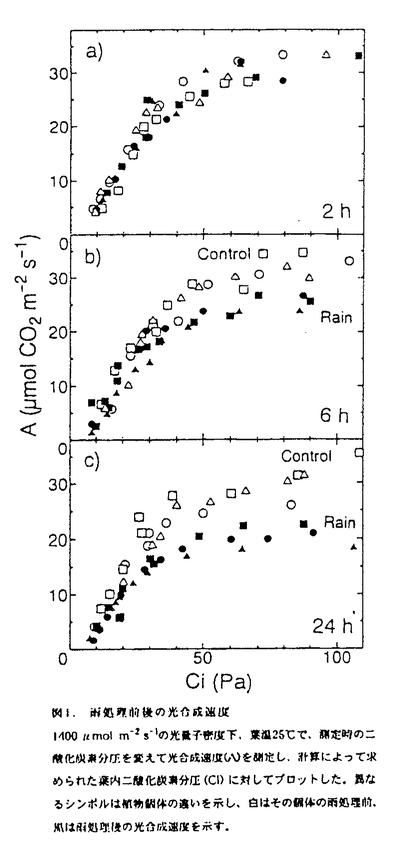
 mol m-2s-1の光量子密度下、葉温25℃で、測定時の二酸化炭素分圧を変えて光合成速度(A)を側定し、計算によって求められた葉内二酸化炭素分圧(Ci)に対してプロットした.異なるシンボルは植物個体の違いを示し、白はその個体の雨処理前.風は雨処理後の光合成速度を示す.
mol m-2s-1の光量子密度下、葉温25℃で、測定時の二酸化炭素分圧を変えて光合成速度(A)を側定し、計算によって求められた葉内二酸化炭素分圧(Ci)に対してプロットした.異なるシンボルは植物個体の違いを示し、白はその個体の雨処理前.風は雨処理後の光合成速度を示す.