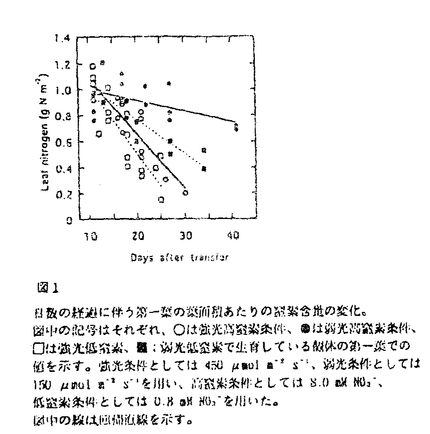
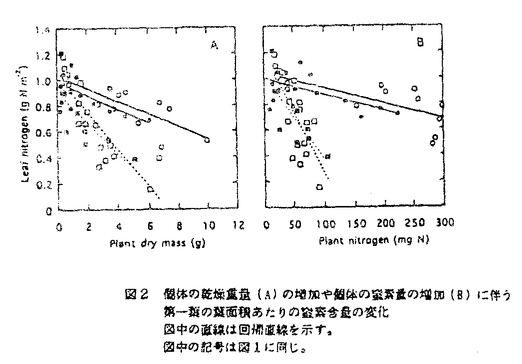
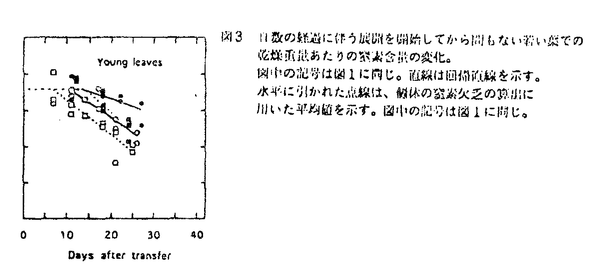
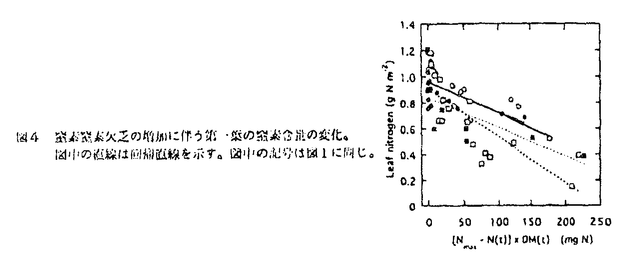
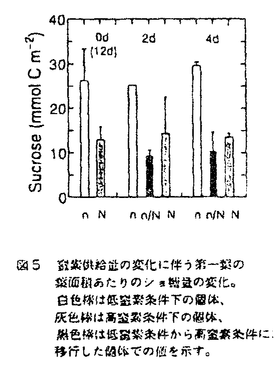
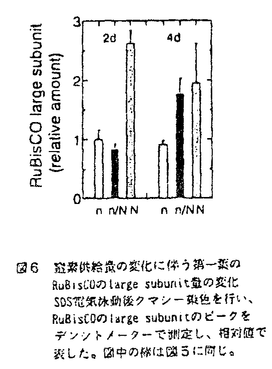
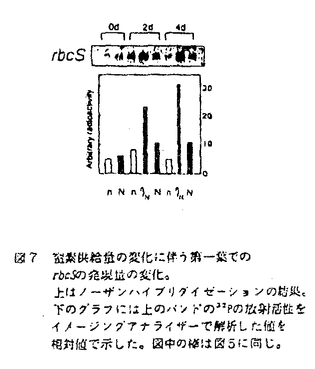
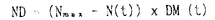本論文は、生物にとって重要な資源である炭素や窒素を有機化する植物が、より効率の良い物質生産のために個体の中でそれらをどのように利用しているのか、また、どのような機構を通して炭素と窒素の利用が制御されているのかという点について、3章に分けて述べている。 第1章では、個葉の老化に伴う窒素量の減少の速さと生育環境との関連、第2章では、個葉に蓄積した光合成産物による光合成系タンパク質をコードする遺伝子発現の抑制、第3章では、個体に様々な被陰処理を行ったときに見られる個葉のタンパク質量の変化について述べている。 葉が展開すると、その葉の窒素量は次第に減少していくが、この減少の速さは、個体のおかれた光環境や、個体への窒素供給量に依存している。これまで、光や窒素の影響は個別に調べられることが多かったが、本論文の第1章では、個体の成長(乾燥重量)と展開し始めた若い葉における窒素量の減少から算出した"個体の窒素欠乏"という新たなパラメータを考案した。光の強度と窒素供給量の影響の両方を含むこのパラメータを導入することにより、これまで報告されてきた個葉の窒素量の減少を統一的に説明できることを明らかにしている。 この"個体の窒素欠乏"の増加が早く、第一葉の窒素量の減少も早い条件下で生育した個体では、第一葉により多くの光合成産物が蓄積するという結果が得られ、第2章では、この光合成産物の蓄積と個葉の窒素量の減少の間に働いている機構を調べている。この章では、個体への窒素供給量を変えることと、光合成阻害剤を用いることによって、老化する個葉に蓄積する糖のレベルの変化と、それに関連する光合成系タンパク質およびその遺伝子の発現レベルの変化を調べている。この章の結果は、切り葉やプロトプラストを用いた系によって明らかにされてきた糖による遺伝子発現の抑制機構が、内生的に光合成産物を蓄積した葉でも働いていることを示唆している。さらに、窒素欠乏条件においてRuBP carboxylase/oxygenase(RuBisCO)量の減少が速い原因として、遺伝子発現の制御を通したその合成の過程に、光合成産物による終産物阻害が働いている可能性を示している。 第3章では、第一葉より上位の葉の光環境の変化が、第一葉のRuBisCO量の減少の速さに与える影響を調べている。この章では、上部が被陰されている場合には、第一葉のおかれた光条件が同じであるにも関わらず、老化に伴うRuBisCO量の減少が遅れるということが示されている。この結果は、上位葉の光環境の変化が何らかの機構を通して第一葉に伝えられ、第一葉のRuBisCO量を変えていることを示唆する。このようなシグナルが何であるかは非常に興味深い問題であり、これを明らかにすることが今後に残された課題である。 本論文は、植物個体の物質生産という生態学的視点から、生理学的手法を用い、系統的に解析を行っている。このような研究方法は、自然界での様々な植物の生き様を明らかにしていく上できわめて重要であると考えられる。 なお、本論文第1章は、寺島一郎、渡邊昭との、第2、3章は渡邊昭との共同研究であるが、論文提出者が主体となって分析及び検証を行ったもので、論文提出者の寄与が十分であると判断する。 よって、博士(理学)の学位を授与できると認める。 |  mol m-2s-1、弱光約150
mol m-2s-1、弱光約150