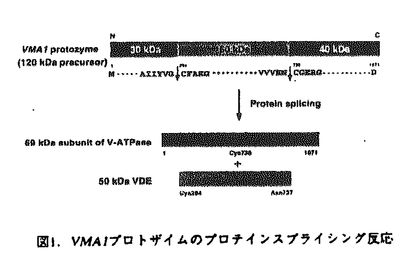
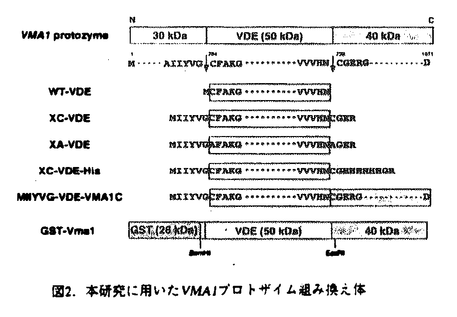
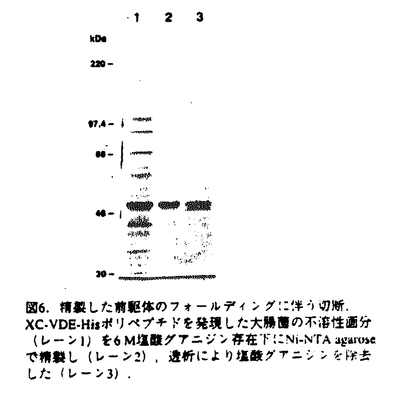
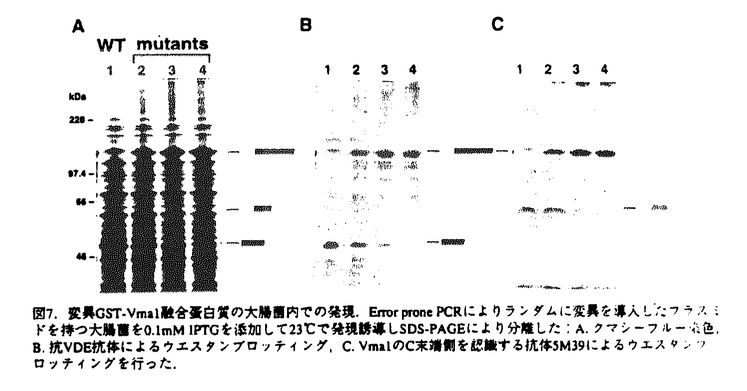
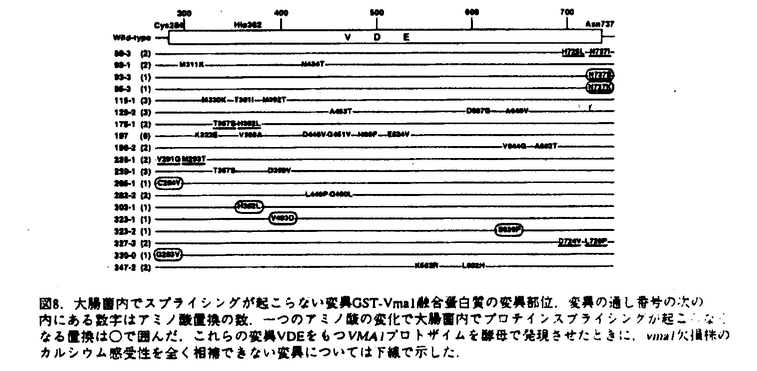
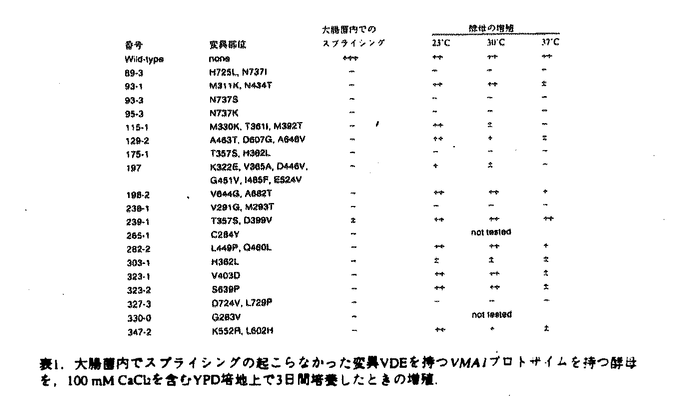
プロテインスプライシングは、1990年,安楽研究室(東大・理)により発見された蛋白質レベルでのスプライシング(ペプチド組継ぎ反応)である。出芽酵母の液胞膜H+-ATPaseの69kDaサブユニットをコードするVMA1遺伝子の中央部分に介在配列が存在し、まず介在配列を含んだまま120kDaの一本の前駆体ポリペプチド(Vma1プロトザイム)として翻訳される。その後に前後の境界部分でペプチド結合の切断が起こって介在配列は切り出され、両端が連結することにより液胞膜H+-ATPaseの本来の69kDaサブユニットが生成する。この切り出される50kDaの介在配列VDE(VMA1-Derived Endonuclease)は、VDEを持たないVMA1遺伝子を特異的に認識して切断するエンドヌクレアーゼ活性を持っており、減数分裂時特異的に働いて自らの遺伝子をVMA1遺伝子に入り込ませる遺伝子ホーミングを触媒する。すなわちVDEは利己的な可動遺伝因子としての性質を持っている。現在までに真核生物、原核生物、古細菌にわたる6つの生物種でプロテインスプライシングの例が報告されている。プロテインスプライシング反応において、切断部位の近傍のアミノ酸が重要な役割を果たしていることが、部位特異的変異導入を用いたin vivoでの解析から明らかとなってきた。 プロテインスプライシング反応はin vivoでは非常に効率よく起こるために、前駆体を単離して生化学的に解析することが困難であった。本研究において、論文提出者,川崎は、プロトザイム組み換え体を大腸菌内で過剰発現させると前駆体がインクルージョンボディに蓄積することを利用し、プロトザイムのフォールディングに依存したin vitroプロテインスプライシング反応系を構築した。この系を利用して、数アミノ酸しか外側に持たないVDEでさえもスプライシング反応が自己触媒的に起こることを見出した。さらに、スプライシング反応に関与するVDE内の一次構造上の領域について変異VDEを用いた解析を行った。研究成果の要旨は以下のとおりである。 1.1.フォールディングに依存したin vitroプロテインスプライシング反応系の構築 プロテインスプライシングにおけるVDEの自己触媒能を調べるために以下の三種類のポリペプチドを大腸菌内で発現させた。WT-VDEはVDE部分に開始Metをつけたもの。XC-VDEはVDEのN末端側に6アミノ酸、C末端側に4アミノ酸をのばしたものである。XA-VDEは切断部位のCysを二つともAlaに換えたもので、切断の起こらないコントロールとして作成した。XC-VDEを発現した大腸菌では、分子量およそ49kDa付近にそれぞれWT-VDE、XA-VDE産物とほぼ同じ移動度を持つ二つのバンドが観察された。このXC-VDEを発現した大腸菌を破壊して遠心すると下のバンドは可溶性画分に、上のバンドは不溶性画分に分離した。すなわち下のバンドは大腸菌内で正しくフォールディングして切断の起こったVDEと考えられ、上のバンドは正しくフォールディングせずに凝集して沈殿し、インクルージョンボディーを形成していると考えられた。 同人は,この未切断のポリペプチドをフォールディングさせることによりプロテインスプライシングをin vitroで行わせることが可能ではないかと予想した。そこで沈殿している未切断のXC-VDEを塩酸グアニジンで変性し可溶化させた後に、透析により塩酸グアニジンを除去してフォールディングさせると、約半数のXC-VDEが切断を受けることを見出した。この透析後のサンプルを遠心すると切断の起こったものは可溶性画分に、切断が起こらなかったものは再び沈殿して不溶性画分に分かれた。そこでさらに、この沈殿を再び塩酸グアニジンで変性し、透析すると、1回目と同様の効率で切断が起こった。 スプライシング部位の化学構造を確定するため,MIIYVG-VDE-VMA1Cを大腸菌内で発現させ、不溶性画分に蓄積した前駆体を変性剤存在下に部分精製し、フォールディングさせた。出現した反応生成物のN末端アミノ酸配列を決定した結果、スプライシングを受けたVDEの両端のペプチドが正しく連結していること,すなわち,VMA1遺伝子は両蛋白質に対する遺伝情報を過不足なく保持していることが確認できた。 このin vitroプロテインスプライシング反応において、大腸菌の不溶性画分に存在する他の成分の影響を除くためにVDEのN末端に6アミノ酸、C末端に6×Hisタグを含む10アミノ酸を付加したポリペプチド(XC-VDE-His)を発現させ、不溶性画分に存在する前駆体ポリペプチドを変性条件下でNi-NTA agaroseを用いて精製した。透析により変性剤を除去すると精製前と同様に効率よく切断が起こったことから、フォールディングに伴うペプチド切断は自己触媒的であることが確認された。前駆体ポリペプチドのフォールディングの条件を検討したところ、切断は中性付近で最も効率よく起こり、また、温度が低いほど効率よく切断が起こった。これは低温ほどフォールディングの時に凝集が起こりにくいためと考えられる。切断は透析による変性剤の除去とともに速やかに進行し、2時間以内に終了した。このフォールディングに伴うin vitroでのペプチドの切断がペプチダーゼ阻害剤により阻害されるか調べたが、通常のペプチダーゼの場合に阻害が見られる濃度ではまったく切断は影響を受けなかった。切断反応はVDE部分の立体構造の形成と同時に非常に効率よく起こり、その反応過程では通常のペプチダーゼ反応における阻害剤が作用する余地のないことを示唆している。さらに、VDEの外側のアミノ酸の長さの異なる2種類の前駆体ポリペプチドを混ぜてフォールディングさせてみたが、スプライシングが2分子間で起こってN末端側とC末端側がつながったものは見出されなかった。これはスプライシング反応が1分子内で起こる反応であることを示唆している。 以上の結果により、VDEはプロテインスプライシング反応によって外側の数アミノ酸のペプチドさえも切断して離脱することができ、それ以外の細胞質因子を何等必要としないこと、すなわちVDEはスプライシング反応を自触的に触媒することが証明された。 2.プロテインスプライシング反応に関わるVDEの一次構造上の要因の解析 これまでに,VDEのN末端側とC末端側の境界部分がスプライシング反応をこ関与することが部位特異的変異導入によりわかっていたが、VDE内部についての情報は無かった。そこでスプライシングにおけるVDE内部の領域の関与を調べるために、スプライシング反応が欠損したVDE領域内の突然変異の単離を行った。 大腸菌内でのスプライシング反応を容易に検出するために、VDEの前にGST、後ろにはVma1のC末端側40kDa部分を持つポリペプチドを発現させた(GST-Vma1)。このGST-Vma1融合蛋白質を大腸菌内で発現させると115kDaのスプライシング前駆体の他に、切り出された50kDa VDEがSDS-PAGE後のクマシーブルー染色により容易に検出される。この融合蛋白質を発現するプラスミドのVDE領域全体を含むBamHI-EcoRI断片をerror prone PCR法により増幅し、ランダムに変異を導入した。324個のコロニーについて調べたところ、大腸菌内でのスプライシングが様々な程度に低下していることが分かった。これらの中で抗VDE抗体によるウェスタンブロッテイングによっても切り出された50kDa VDEが検出されず、スプライシング前駆体が蓄積している17種類の変異プラスミドについてそのDNA配列を解析し、変異部位を決定した。 次に、大腸菌を用いたスクリーニングにより得られたこれらのスプライシング反応欠損変異VDEをVma1プロトザイムに導入したときに、酵母の中でもスプライシングが起こらないかどうか調べた。スプライシングの有無をvma欠損株の示す特徴的な表現型であるカルシウム感受性を指標に調べたところ、いくつかの変異でvma1欠損を部分的に相補することができた。これらの変異Vma1プロトザイムが酵母内で低い効率ながらスプライシングを行うことはウェスタンプロッティングによっても確認された。したがって変異によるダメージは、酵母内におけるよりも、大腸菌内でより強く表れたと考えられる。酵母内でもスプライシングが全く起こらない6つの変異VDEについては、特に反応に重大な欠損があると考えられる。これらの変異は3つの領域、すなわちN末端およびC末端側の2つの境界部分とVDE内部のHis362を中心とする領域に分布していた。以上の結果から、これまで指摘されていたN末端とC末端の境界部分に加えて、VDE内部のHis362残基周辺の領域が特にプロテインスプライシング反応に関与することが示された。一方、これらの領域に変異を持つVDEは、エンドヌクレアーゼ活性は正常であった。したがってVDEの持つ2つの機能(プロテインスプライシングとエンドヌクレアーゼ)は独立していることが示唆された。 本研究により,プロテインスプライシング反応のin vitro検出系が確立され,これを利用した分子生物学的・生化学的研究から,本反応の自触的反応特性と組継ぎ機構の分子的理解が確立された.よって,審査委員は,全員一致して,この研究が博士(理学)の学位に充分な業績であると認定した. |