脳下垂体前葉から分泌される生殖腺刺激ホルモンには濾胞刺激ホルモン(FSH)と黄体形成ホルモン(LH)があり、いずれも分子量約3万の糖蛋白質である。両ホルモンとも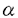 、 、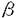 と名づけられた2つのサブユニットが結合したヘテロダイマーであり、サブユニットはTSHも含めた脳下垂体の糖蛋白質ホルモンに共通のものだが、サブユニットは各ホルモンに固有でホルモンの特異的な機能はこの部分の構造による。また、FSHとLHはそれぞれに特異的な受容体分子と結合することでその機能を発揮する。FSHは卵巣濾胞の成熟をもたらし、精巣ではアンドロゲンと共に精子形成を促す。LHは精巣ライディヒ細胞のアンドロゲン産生を促し、卵巣に対しては濾胞のエストロゲン産生を刺激し、さらに排卵を引き起こす。FSH受容体(FSH-R)とLH受容体(LH-R)のクローニングは哺乳類でのみ報告されており、両受容体とも7回細胞膜を貫通するG蛋白質結合型の糖蛋白質である。FSH-R分子は365-366アミノ酸残基の細胞膜外領域、264アミノ酸残基の膜貫通領域、63-65アミノ酸残基の細胞膜内領域からなり、LH-R分子は333-335残基の細胞膜外領域、266-267残基の膜貫通領域、70-72残基の細胞内領域からなる。FSH-RおよびLH-Rともに他の7回膜貫通型の受容体に比べ、細胞膜外領域が大きいのが特徴である。アミノ酸レベルでのFSH-RとLH-Rの相同性は膜貫通領域で比較すると約80%にもなるが、細胞膜外領域では50%程度にすぎず、合成蛋白質を用いた研究からホルモンとの結合の特異性は細胞膜外領域によって担われていることが明らかにされている。しかし、比較的相同性の低い細胞膜外領域でもFSH-RとLH-Rに共通した特定の残基の繰り返し構造が見出されていることから、これらの遺伝子は共通の祖先分子から進化してきたと考えられている。このため細胞膜外領域にはホルモンとの結合の選択性だけでなく、分子進化の足跡となる鍵が記されていると考えられる。そこで私は、哺乳類以外の脊椎動物の生殖腺刺激ホルモン受容体についてホルモン結合部位の構造を比較検討し分子進化の一端を探り、さらにこの領域の配列上の特異性を利用して胚生殖腺でのmRNAの発現開始と局在を明らかにし、性的二形の成立機構についての理解を深めることを研究の目的とした。 と名づけられた2つのサブユニットが結合したヘテロダイマーであり、サブユニットはTSHも含めた脳下垂体の糖蛋白質ホルモンに共通のものだが、サブユニットは各ホルモンに固有でホルモンの特異的な機能はこの部分の構造による。また、FSHとLHはそれぞれに特異的な受容体分子と結合することでその機能を発揮する。FSHは卵巣濾胞の成熟をもたらし、精巣ではアンドロゲンと共に精子形成を促す。LHは精巣ライディヒ細胞のアンドロゲン産生を促し、卵巣に対しては濾胞のエストロゲン産生を刺激し、さらに排卵を引き起こす。FSH受容体(FSH-R)とLH受容体(LH-R)のクローニングは哺乳類でのみ報告されており、両受容体とも7回細胞膜を貫通するG蛋白質結合型の糖蛋白質である。FSH-R分子は365-366アミノ酸残基の細胞膜外領域、264アミノ酸残基の膜貫通領域、63-65アミノ酸残基の細胞膜内領域からなり、LH-R分子は333-335残基の細胞膜外領域、266-267残基の膜貫通領域、70-72残基の細胞内領域からなる。FSH-RおよびLH-Rともに他の7回膜貫通型の受容体に比べ、細胞膜外領域が大きいのが特徴である。アミノ酸レベルでのFSH-RとLH-Rの相同性は膜貫通領域で比較すると約80%にもなるが、細胞膜外領域では50%程度にすぎず、合成蛋白質を用いた研究からホルモンとの結合の特異性は細胞膜外領域によって担われていることが明らかにされている。しかし、比較的相同性の低い細胞膜外領域でもFSH-RとLH-Rに共通した特定の残基の繰り返し構造が見出されていることから、これらの遺伝子は共通の祖先分子から進化してきたと考えられている。このため細胞膜外領域にはホルモンとの結合の選択性だけでなく、分子進化の足跡となる鍵が記されていると考えられる。そこで私は、哺乳類以外の脊椎動物の生殖腺刺激ホルモン受容体についてホルモン結合部位の構造を比較検討し分子進化の一端を探り、さらにこの領域の配列上の特異性を利用して胚生殖腺でのmRNAの発現開始と局在を明らかにし、性的二形の成立機構についての理解を深めることを研究の目的とした。 まず始めに各種脊椎動物の生殖腺刺激ホルモン受容体のホルモン結合部位を単離し、分子構造を明らかにするためRT-PCRクローニングを行なった。その結果、鳥類(ウズラ)のLH-RとFSH-Rのホルモン結合部位の一次構造が明らかになった。特にFSH-Rに関しては3種の爬虫類(クサガメ、ヤモリ、トカゲ)の一次構造も明らかにしたので、現生有羊膜類に至る三つの大きなグループ、すなわち無弓亜綱(カメにいたる)、単弓亜綱(哺乳類にいたる)、双弓亜綱(鳥類、ワニ、トカゲに至る)の3系統のデータが得られたことになる。その結果、ロイシン、イソロイシン、ヴァリンおよびフェニルアラニンからなる繰り返しのモチーフは、すべての分子で保存されていることが明らかになった。このため一次構造上の相同性は高く、哺乳類のものと比較した場合、アミノ酸レベルで約65%であった(ラットLH-R-ウズラLH-R,67%;ラットFSH-R-ウズラFSH-R,67%;ラットFSH-R-クサガメFSH-R,70%;ラットFSH-R-ヤモリFSH-R,57%;ラットFSH-R-トカゲFSH-R,68%)。従来のバインディングアッセイにより、LHはFSHよりも種特異性が高いことが明らかにされてきている。一次構造の上でもFSHサブユニットはウズラと哺乳類とで64-69%の相同性をもち、LHサブユニットでは約50%であるから、FSHの保守性はLHよりも高いといえる。しかし受容体については、FSH-RおよびLH-Rともにウズラと哺乳類間の相同性はほぼ等しかった。個々の残基に注目した場合、糖鎖修飾の推定される部分およびシステイン残基の位置にいくつかの差異が見出された。哺乳類では糖鎖修飾の予想される部位は3個所あり、これまでに報告されているすべての種で保存されている。ところがウズラとヤモリではこのうち1個所を欠き、異なる部位にウズラ、クサガメ、ヤモリで1個所、トカゲで2個所見出された。ラットを用いた研究から糖鎖修飾を正しく受けることは、FSH-Rが正しく細胞膜上に発現するために必須であることが示されている。従って解析したすべての種で保存されていた2個所の修飾部位が特に重要と思われる。細胞膜外領域のシステイン残基はS-S結合の形成などを通じて、蛋白質の立体構造の形成に重要であると考えられる。事実、哺乳類では6個所ある残基がすべての種で保存されている。また、ウズラではそのうち一つの位置が異なるのみであった。他方、爬虫類においては、そのうちの1個所(ラットCys188に相当する残基)が失われていた。さらにヤモリにおいては、他種に見られない位置にシステイン残基が存在していた。この他、FSH受容体のエクソン10(最後のエクソン)のN’端側に該当する部分に、際立って相同性の低い部分が見出された。この部位において、クサガメではラットに比較して3残基多く、ヤモリは7残基、トカゲは19残基少なかった。しかしながら推定されるアミノ酸配列の疎水性を解析したところ、この領域は親水性の高い部分であるという特性は保存されていた。 続いて本研究で明らかにした配列をデータとして各分子間の系統関係を考察した。平均距離法(UPGMA)、近隣結合法(NJ)、最大節約法(MP)の3法を用いて分子系統樹を構築したところ、いずれの方法でもヤモリFSH-RはトカゲFSH-Rとクラスターを形成した。クサガメFSH-RはUPGMAとNJではウズラFSH-Rとクラスターしたが、MPではヤモリ-トカゲのクラスターの枝とクラスターを形成した(従来の一般的な種系統樹にはMPによる結果の方が近い)。いずれの方法でも哺乳類(単弓亜綱)の枝と鳥類・爬虫類の枝は最初に分かれており、ラットCys188に相当するシステイン残基は哺乳類にいたる系統の途中で出現したか、あるいは無弓・双弓亜綱に至る経路の途中で失われたものらしい。また有鱗類(ヤモリ、トカゲ)に特異的なエクソン10におけるアミノ酸残基の欠失は、双弓亜綱の中で有鱗類が別れた三畳紀以降に生じたものと推定した。 さらに本研究でニワトリ胚の生殖腺におけるLH-R mRNAの発現を解析した。LH-Rの発現は生殖腺の発育、ステロイドホルモンの合成に必須の要素である。特に胚発生の初期におけるステロイドホルモン合成開始に重要であるが、これまでに鳥類ではLHとステロイドホルモンの免疫組織学的な検出報告があるのみであった。 初期胚の生殖腺原基は精巣・卵巣のいずれにも分化していない。ニワトリの場合、孵卵6日目までは形態的に雌雄の区別はつかないが、7日目ころから精巣と卵巣に違いがはっきりしてくる。すなわち精巣・卵巣とも当初髄索が発達するが、この頃から卵巣では左側のみ皮索が発達しはじめ、髄索は退縮しはじめる。LH-RのcRNAをプローブとしたin situハイブリダイゼーションをニワトリ胚(5日胚-孵化後1日)に対して行なったところ、卵巣では早いものでは孵卵7日から、精巣では孵卵14日目からシグナルが検出された。これらの結果は血中と脳下垂体中のLHの存在に関する従来の知見と整合するものである。ニワトリの孵化に要する孵卵日数は21日であり、ラットの妊娠期間と同じである。ラットの場合LH-R mRNAの発現は精巣で胎齢の14.5日目から、卵巣では17日目から(いずれもRT-PCRによる検出)と報告されており、鳥類のLH-R発現は哺乳類よりもかなり早いといえる。特にニワトリ卵巣のLH-R mRNA発現開始は精巣に比較して約1週間早かった。メスヘテロ型の性決定様式に従う鳥類の生殖腺とその付属器官の原基の形態はオスが基本型である。鳥類がメスらしさを獲得するために胚発生時にいち早く、卵巣がLHに応答しエストロゲンを分泌することはきわめて重要なのであろう。なおニワトリの卵巣は左側のみ発育し、右側卵巣は孵卵10日目ころから退化し始める。本研究では、この右側卵巣においても少なくとも孵化するまでLH-R mRNAの発現が検出された。生殖腺刺激ホルモンがなぜ右側卵巣を維持できないのかは今後の課題である。 精巣におけるLH-R mRNAの発現は細精管の間を埋める間質に局在していた。間質細胞は将来ライディッヒ細胞に分化するものと考えられ、成体(ウズラ)での結果と合致するものである。一方、卵巣のLH-R mRNA発現の場は孵卵中を通して髄索であった。胚卵巣におけるステロイド合成関連の諸酵素は髄索に局在するので、この結果は妥当といえる。ただし髄索は卵巣の発生途上で退化してゆくもので、成体におけるLHの標的細胞は顆粒膜細胞、莢膜細胞および間質細胞であり、これらは皮索から発達するとされている。従って胚と成体とではLH-R mRNAを発現する細胞の起源が異なっていると思われ、孵化後性的に成熟するにしたがいLHの標的細胞は上記の各細胞に移ってゆくと考えられる。 本研究により哺乳類以外の脊椎動物の生殖腺刺激ホルモン受容体の一次構造を明らかにし、この結果に基づき分子間の系統関係を考察した。またニワトリ胚において、性的二形の成立に関連するLH-R mRNAの発現の開始時期と局在を明らかにした。 |