| 内容要旨 | | 変態は幼生から成体への移行の際におこる体制の大きな変換を含む形態形成の過程の一つであり、様々な動物でその機構が調べられてきた。ホヤはその変化が他の実験動物と比べて短いこと、幼生を様々に環境を変化させた海水で飼育し、生理学的な実験を行なうことができることなどから変態機構を研究するのに適している。そこで、ホヤ幼生を研究材料に用いて付着突起の崩壊と尾部吸収の機構とそれ以降の形態変化及びそれらの関連性についての研究を行った。 1.カタユウレイボヤ幼生の変態過程における付着突起の形態変化、尾部吸収、胴体の分化についての形態学的、生理学的解析 ホヤ幼生はフ化後遊泳生活を始め、しばらくして胴部先端にある付着突起で基板に付着し、付着突起が形態変化をおこし、次いで幼生の尾部が一気に吸収され、最後に体軸が回転をし成体へと変態する。これまで付着、尾部吸収がほとんどの場合この順序で起こるため、付着突起は刺激の受容器官であり、付着突起で受け取った刺激をシグナルとしてその後の変態が起こると考えられてきた。本研究ではまず、カタユウレイボヤ遊泳幼生の変態過程を自然な状態で詳細に観察したところ、従来の報告とは異なり、付着前の遊泳期に付着突起の形態変化が起こっていること(図1-1)、尾部吸収は、付着後、付着突起の崩壊前に尾部の後端からはじまり、付着突起が崩壊する頃、尾部の胴体側での吸収が始まること(図1-2)を明らかにした。このことは、正常発生において、付着突起の崩壊の過程とはじめの尾部吸収の過程は、異なるタイミングで起きていることを示している。次に、付着前におこる付着突起の形態変化及び付着を、低温処理、またはカルシウムのキレーター試薬処理で抑制し、遊泳期における付着突起の形態変化と尾部吸収の関係を調べたところ、付着及び付着突起の形態変化を伴なわない尾部吸収が起こることを明らかにした(図-2)。この時起こった尾部吸収は部分的であることから、尾部吸収はこの部分吸収及び付着突起の崩壊を伴なう完全な吸収の二段階に分けられると考えられる。付着突起で付着突起の形態変化に反映されない変態へのシグナルを付着突起で受け取っている可能性を除去するために、機械的に幼生を切断し、付着突起から尾部への仮想的な刺激経路を切断したところ、切断時期、部域に関わらず正常発生の幼生とほぼ同時期に尾部吸収がはじまり、胴体断片は尾部無しでも体軸が回転し成体器官の分化が起こった(図-3)。また、脳胞を含む尾部断片では、尾部吸収が完全に終了したが、含まない断片では完全には終了しなかった。さらに、脳胞を含む尾部断片ではテスト細胞の移動は正常に起こり(パート2参照)、それに伴う被嚢、表皮における多糖の変化も正常発生の幼生と同様な変化を示したが(パート3参照)、脳胞を含まない尾部断片ではテスト細胞の移動も多糖の変化も正常発生の幼生とは異なった(図-4)。 以上の結果から、カタユウレイボヤでは従来の仮説のように変態過程の全てが付着、尾部吸収、成体器官の分化の順のシグナルトランスダクション機構によって起こっているのではなく、この連続機構と各々の独立した機構の組み合わせによって変態が進行すると考えられる。 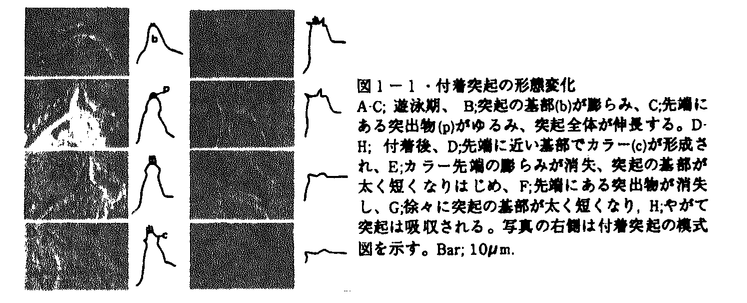 図1-1・付着突起の形態変化A-C;遊泳期、B;突起の基部(b)が膨らみ、C;先端にある突出物(p)がゆるみ、突起全体が伸長する。D-H;付着後、D;先端に近い基部でカラー(c)が形成され、E;カラー先端の膨らみが消失、突起の基部が太く短くなりはじめ、F;先端にある突出物が消失し、G;徐々に突起の基部が太く短くなり,H;やがて突起は吸収される。写真の右側は付着突起の模式図を示す。Bar;10 図1-1・付着突起の形態変化A-C;遊泳期、B;突起の基部(b)が膨らみ、C;先端にある突出物(p)がゆるみ、突起全体が伸長する。D-H;付着後、D;先端に近い基部でカラー(c)が形成され、E;カラー先端の膨らみが消失、突起の基部が太く短くなりはじめ、F;先端にある突出物が消失し、G;徐々に突起の基部が太く短くなり,H;やがて突起は吸収される。写真の右側は付着突起の模式図を示す。Bar;10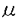 m. m. 図1-2・尾部の形態変化A;付着直後の幼生、付着突起(▼)は、形態的に付着前と差はまだ見られないが、尾部の後端(↓)ではすでに吸収が始まっている。B;付着後6-7hr、尾部前端(胴体)脊索(nt)で吸収に伴う折れ曲がりが見られる。この時、付着突起(▼)は崩壊している。Bar;10m. 図1-2・尾部の形態変化A;付着直後の幼生、付着突起(▼)は、形態的に付着前と差はまだ見られないが、尾部の後端(↓)ではすでに吸収が始まっている。B;付着後6-7hr、尾部前端(胴体)脊索(nt)で吸収に伴う折れ曲がりが見られる。この時、付着突起(▼)は崩壊している。Bar;10m. 図2・低温処理の変態への影響A,B;低温処理中の全体像と付着突起。A;尾部後端(▼)、及び前端での吸収が始まっている。B;付着突起(▼)は図1-1Aの形態を保っている。C,D;常温へ戻した時の全体像と付着突起。D;付着突起(▼)は崩壊する。C;脊策の空胞が無くなり、尾部吸収は一気に進む。Bar;A,C;50m,B,D;10m. 図2・低温処理の変態への影響A,B;低温処理中の全体像と付着突起。A;尾部後端(▼)、及び前端での吸収が始まっている。B;付着突起(▼)は図1-1Aの形態を保っている。C,D;常温へ戻した時の全体像と付着突起。D;付着突起(▼)は崩壊する。C;脊策の空胞が無くなり、尾部吸収は一気に進む。Bar;A,C;50m,B,D;10m.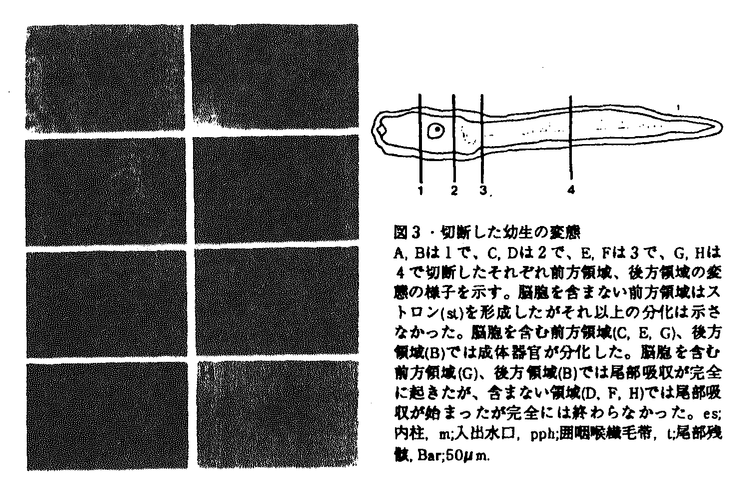 図3・切断した幼生の変態A,Bは1で、C,Dは2で、E,Fは3で、G,Hは4で切断したそれぞれ前方領域、後方領域の変態の様子を示す。脳胞を含まない前方領域はストロン(st)を形成したがそれ以上の分化は示さなかった。脳胞を含む前方領域(C,E,G)、後方領域(B)では成体器官が分化した。脳胞を含む前方領域(G)、後方領域(B)では尾部吸収が完全に起きたが、含まない領域(D,F,H)では尾部吸収が始まったが完全には終わらなかった。es;内柱,m;入出水口,pph;囲咽喉繊毛帯,t;尾部残骸,Bar;50m.2.遊泳期幼生におけるテスト細胞の移動と被嚢形成 図3・切断した幼生の変態A,Bは1で、C,Dは2で、E,Fは3で、G,Hは4で切断したそれぞれ前方領域、後方領域の変態の様子を示す。脳胞を含まない前方領域はストロン(st)を形成したがそれ以上の分化は示さなかった。脳胞を含む前方領域(C,E,G)、後方領域(B)では成体器官が分化した。脳胞を含む前方領域(G)、後方領域(B)では尾部吸収が完全に起きたが、含まない領域(D,F,H)では尾部吸収が始まったが完全には終わらなかった。es;内柱,m;入出水口,pph;囲咽喉繊毛帯,t;尾部残骸,Bar;50m.2.遊泳期幼生におけるテスト細胞の移動と被嚢形成 幼生の被嚢の外側にあるテスト細胞は従来極めて静的な存在であると考えられてきた。本研究では、変態に先立つ遊泳期の幼生について光学顕微鏡、透過型電子顕微鏡を用いて詳細な観察を行ない、その形態的な特徴から、遊泳期の幼生を3ステージに分けた。ステージ1(孵化後0-3hr,20C゜)では、胴体で、成体被嚢(C2)が幼生被嚢(C1)と表皮の間に形成されはじめ、テスト細胞は幼生被嚢の外側に均一に分布している状態を示す。ステージ2(孵化後3-5hr,20C゜)では、胴体でC1とC2が膨らみはじめ、テスト細胞はC1上で移動を開始する。ステージ3(孵化後5-9hr,20C゜)では、胴体でC1とC2が尾部の方へ引かれるように二つの山型を形成し、テスト細胞は、胴体で6条の線上に並び、尾部では表皮に沿った頭尾軸上に並ぶ(図-4)。電顕の観察から、テスト細胞は尾部で、ステージ3の時期に、被嚢の中へ侵入し、表皮から伸びる感覚毛に接触することを明らかにした(図-4)。また、テスト細胞は、その後付着直前に被嚢上へ分散した(パート1参照)。以上のテスト細胞の時期部域特異的な挙動から、テスト細胞の変態への関与が示唆された。 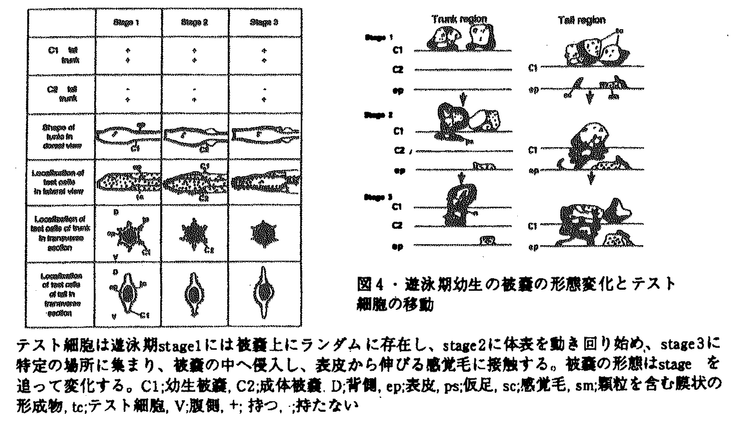 図4・遊泳期幼生の被嚢の形態変化とテスト細胞の移動テスト細胞は遊泳期stage1には被嚢上にランダムに存在し、stage2に体表を動き回り始め、stage3に特定の場所に集まり、被嚢の中へ侵入し、表皮から伸びる感覚毛に接触する。被嚢の形態はstageを追って変化する。C1;幼生被嚢,C2;成体被嚢.D;背側,ep;表皮,ps;仮足,sc;感覚毛,sm;顆粒を含む膜状の形成物,tc;テスト細胞,V;腹側,+;持つ,;持たない3.テスト細胞を除去した幼生におけるテスト細胞様の新しい細胞の新生と正常な変態過程の撹乱 図4・遊泳期幼生の被嚢の形態変化とテスト細胞の移動テスト細胞は遊泳期stage1には被嚢上にランダムに存在し、stage2に体表を動き回り始め、stage3に特定の場所に集まり、被嚢の中へ侵入し、表皮から伸びる感覚毛に接触する。被嚢の形態はstageを追って変化する。C1;幼生被嚢,C2;成体被嚢.D;背側,ep;表皮,ps;仮足,sc;感覚毛,sm;顆粒を含む膜状の形成物,tc;テスト細胞,V;腹側,+;持つ,;持たない3.テスト細胞を除去した幼生におけるテスト細胞様の新しい細胞の新生と正常な変態過程の撹乱 受精後、機械的、化学的にテスト細胞を除去し、発生させた幼生を作成した。テスト細胞を除去すると、まず、尾芽胚期に胚の外側にテスト細胞と異なる性質を持つ新しい細胞が出現し、遊泳期間にその数は増減する(図-5)。細胞組織化学的な判定から、この新しい細胞は正常発生では、発生しない細胞であることが明らかとなった。テスト細胞を持たない幼生は、形態的には正常発生の幼生とほとんど差は無いが、正常幼生では変態直前に起こる尾部表皮、被嚢の多糖の変化がおこらない。正常発生の幼生では、変態過程において、付着後、尾部が完全に吸収してから、体軸が回転していくのに対して、テスト細胞を除去した幼生では、尾部が途中まで吸収し、その後体軸が回転してから、残りの尾部が吸収する、つまり、尾部吸収が時期によって二段階に分けられることを示した(図-6)。テスト細胞は二段階目の尾部吸収のタイミングを制御している可能性を示唆している。以上からテスト細胞がホヤ変態過程の調節に大きく関与していることが明らかとなった。  図5・テスト細胞を除去した胚における新しい細胞の出現A;テスト細胞を除去した胚では尾芽胚前期までは、胚外に細胞は見られない。B;尾芽胚後期になるとまず、胴体と尾部のつけね付近に新しい細胞(▼)が現われる。C;その数は増加し、遊泳期中に増減する。Bar;50m. 図5・テスト細胞を除去した胚における新しい細胞の出現A;テスト細胞を除去した胚では尾芽胚前期までは、胚外に細胞は見られない。B;尾芽胚後期になるとまず、胴体と尾部のつけね付近に新しい細胞(▼)が現われる。C;その数は増加し、遊泳期中に増減する。Bar;50m. 図6・テスト細胞除去卵由来の幼生の変態過程A-C;正常発生幼生の変態過程,D-F;テスト細胞除去卵由来の幼生の変態過程,A;正常発生では幼生は、完全に尾部が吸収した後、B;体軸が回転し、C;様々な成体器官を分化させ、入出水口(m)を開口し、節食を始める。D;テスト細胞除去卵由来の幼生では、途中まで尾部が回転した後、体軸が回転し始める。E;尾部吸収途中でさらに体軸は回転し、様々な成体器官が分化し始め、F;体軸の回転が終了した後、残りの尾部が吸収され、入出水口を開口し節食を始める。ep;表皮,es;内柱,gs;鰓裂,int;腸,pph;囲咽喉繊毛帯,s;胃,st;ストロン,ta;尾部,Bar;50m. 図6・テスト細胞除去卵由来の幼生の変態過程A-C;正常発生幼生の変態過程,D-F;テスト細胞除去卵由来の幼生の変態過程,A;正常発生では幼生は、完全に尾部が吸収した後、B;体軸が回転し、C;様々な成体器官を分化させ、入出水口(m)を開口し、節食を始める。D;テスト細胞除去卵由来の幼生では、途中まで尾部が回転した後、体軸が回転し始める。E;尾部吸収途中でさらに体軸は回転し、様々な成体器官が分化し始め、F;体軸の回転が終了した後、残りの尾部が吸収され、入出水口を開口し節食を始める。ep;表皮,es;内柱,gs;鰓裂,int;腸,pph;囲咽喉繊毛帯,s;胃,st;ストロン,ta;尾部,Bar;50m. (まとめ)ホヤ幼生の変態において、(1)尾部吸収の第一段階は少なくとも、付着突起で受け取る刺激とは無関係に、発生のある段階が来れば自律的に起こること、(2)その後の完全な尾部吸収の終了には、脳胞が重要な役割を果たしていること、(3)そのタイミングの制御にテスト細胞が重要な役割を果たしていることが明らかとなった。 |
| 審査要旨 | | 本論文は3章からなり、第1章はカタユウレイボヤ幼生の変態過程における形態変化について述べられ、又、変態に関わる生理学的解析を行っている。第2章では、テスト細胞のカタユウレイボヤ幼生の遊泳期における移動について述べられ、この細胞の被嚢形成及び変態における役割についての解析を行っている。第3章では実際に受精卵からテスト細胞を除去し、テスト細胞欠如幼生を作成しその形態変化等からテスト細胞が変態において重要な働きを果たしていることが述べられている。 第1章においては次のことが述べられている。ホヤ幼生はフ化後遊泳生活を始め、しばらくして胴部先端にある付着突起で基板に付着し、付着突起が形態変化をおこし、次いで幼生の尾部が一気に吸収され、最後に体軸が回転をし成体へと変態する。これまで付着、尾部吸収がほとんどの場合この順序で起こるため、付着突起は刺激の受容器官であり、付着突起で受け取った刺激をシグナルとしてその後の変態が起こると考えられてきた。本章ではまず、(1)カタユウレイボヤ遊泳幼生の変態過程を自然な状態で詳細に観察したところ、従来の報告とは異なり、付着前の遊泳期に付着突起の形態変化が起こっていること、尾部吸収は、付着後、付着突起の崩壊前に尾部の後端からはじまり、付着突起が崩壊する頃、尾部の胴体側での吸収が始まることを明らかにした。このことは正常発生において、付着突起の崩壊の過程とはじめの尾部吸収の過程は、異なるタイミングで起きていることを示している。次に、(2)付着前におこる付着突起の形態変化及び付着を、低温処理、またはカルシウムのキレーター試薬処理で抑制したところ、付着及び付着突起の形態変化を伴なわない第一段階の尾部吸収と付着突起の崩壊を伴う第2段階の完全な吸収が存在することが明らかになった。更に(3)機械的に幼生を切断し、付着突起から尾部への刺激経路を切断したところ、切断時期、部域に関わらず正常発生の幼生とほぼ同時期に尾部吸収がはじまり、胴体断片は尾部無しでも体軸が回転し成体器官の分化が起こることを示した。また、脳胞を含む尾部断片では、尾部吸収が完全に終了したが、含まない断片では完全には終了しないことを明らかにした。さらに、脳胞を含む尾部断片ではテスト細胞の移動は正常に起こり、それに伴う被嚢、表皮における多糖の変化も正常発生の幼生と同様な変化を示したが、脳胞を含まない尾部断片ではテスト細胞の移動も多糖の変化も正常発生の幼生とは異なることが示された。以上の結果から、カタユウレイボヤでは従来の仮説のように変態過程の全てが付着、尾部吸収、成体器官の分化の順の機構によって起こるのではなく、この連続機構と各々の独立した機構の組み合わせによって変態が進行すること、又、脳胞が変態機構に重要であることが明らかとなった。 第2章において次のことが述べられている。従来幼生の被嚢の外側にあるテスト細胞は極めて静的な存在であると考えられてきた。本研究では、変態に先立つ遊泳期の幼生について光学顕微鏡、透過型電子顕微鏡を用いて詳細な観察を行ない、その形態的な特徴から、遊泳期の幼生を3つのstageに分けた。stage1では、胴体で、成体被嚢(C2)が幼生被嚢(C1)と表皮の間に形成されはじめ、テスト細胞は幼生被嚢の外側に均一に分布している状態を示す。stage2では、胴体でC1とC2が膨らみはじめ、テスト細胞はC1上で移動を開始する。stage3では、胴体でC1とC2が尾部の方へ引かれるように二つの山型を形成し、テスト細胞は胴体で6条の線上に並び、尾部では表皮に沿った頭尾軸上に並ぶことが明らかとなった。更に電顕の観察から、テスト細胞は尾部で、stage3の時期に、被嚢の中へ侵入し、表皮から伸びる感覚毛に接触することを明らかにした。また、テスト細胞は、その後付着直前に被嚢上へ分散した。以上のテスト細胞の時期部域特異的な挙動から、テスト細胞の変態への関与が強く示唆された。 第3章においては次のことが述べられている。受精後、機械的、化学的にテスト細胞を除去し、発生させテスト細胞欠損幼生を作成した。この幼生においては、まず(1)尾芽胚期に胚の外側にテスト細胞と異なる性質を持つ新しい細胞が出現し、遊泳期間にその数は増減する。細胞組織化学的な判定から、この新しい細胞は正常発生では、発生しない細胞であることが明らかとなった。(2)テスト細胞欠損幼生は、形態的には正常発生の幼生とほとんど差は無いが、正常幼生で変態直前に起こる尾部表皮、被嚢の多糖の変化がおこらない。正常発生の幼生では、変態過程において、付着後、尾部が完全に吸収してから、体軸が回転していくのに対して、テスト細胞欠損幼生では、尾部が途中まで吸収し、その後体軸が回転してから、残りの尾部が吸収する、つまり、尾部吸収が時期によって2段階に分けられることが明らかとなった。即ち、テスト細胞は2段階目の尾部吸収のタイミングを制御している可能性を示唆している。以上からテスト細胞がホヤ変態過程の調節に大きく関与していること、及び新しい細胞の出現が明らかとなった。 以上をまとめるとホヤ幼生の変態において、(1)付着突起で受け取る刺激とは無関係に、発生のある段階が来れば自律的に起こる第1段階の尾部吸収、脳胞が重要な役割を果たしている第2段階目の完全な尾部吸収があることが新たに明らかにされた。更に、変態のタイミングの制御にテス細胞が重要な役割を果たしていることが明確となった。 尚、本論文第1章は森沢正昭博士と、第2章は森沢正昭博士、寺門潔博士と、第3章は森沢正昭博士との共同研究であるが論文提出者が主体となって観察、解析、検証等を行ったので論文提出者の寄与が十分であると判断する。よって博士(理学)の学位を授与できると認める。 |