多精を拒否し、受精を正常に行うことは、その後の個体発生を正常に行わせるために必要な前提条件である。多くの種においては、多精拒否機構は二つの段階から成ることが知られている。一つは卵細胞膜の脱分極による早い多精拒否機構であり、一つは表層粒の崩壊(表層反応)、卵外被の構造変化を伴う遅い多精拒否機構である。海産二枚貝卵では、これまでよく研究されてきたウニなどとは異なり、受精後の表層反応・受精膜形成などの形態変化が全く観察されておらず、多精拒否機構も全く不明であった。そこで本研究ではムラサキイガイ及びマガキの卵における多精拒否機構について検討した。 第1部ムラサキイガイ卵の多精拒否機構 Dufresne-Dube et al.(1983)は、Mytilus galloprovincialisの卵に低濃度の精子で媒精しても容易に多精を誘起できることから、Mytilus galloprovincialisの多精拒否機構は不完全であると結論した。しかし本研究第1部においてこの点をムラサキイガイ(Mytilus edulis)で再検討したところ、放卵直後の卵は多精になりにくく、放卵後の時間に依存して多精が誘起されやすくなることを見出した。 次に多精拒否の機構について検討した。ムラサキイガイ卵を人工海水中のNa+濃度を低くして媒精すると多精が誘起されたことから、Na+の流入に伴う脱分極に依存した電気的な早い多精拒否機構が存在すると考えられた。そこで微小電極により卵細胞膜の膜電位変化を測定したところ、受精直後から30秒間脱分極が観察された。更に外液のNa+濃度を低くすると脱分極は小さくなり、多精が誘起された。これらのことからムラサキイガイ卵に電気的な早い多精拒否機構が存在していることが示唆された。 一方、卵細胞膜が脱分極状態から静止電位に戻った時点で、受精卵を高濃度の精子で再び媒精すると、精子侵入が阻害された。このことは電気的な早い機構に続く、遅い多精拒否機構が存在することを示唆している。再媒精した精子の大部分は先体反応を抑制され、卵への結合を阻害されていたが、一部の精子は先体反応を起こして卵へ結合した。再媒精した精子のうち、卵に結合した精子を透過型電子顕微鏡で観察したところ、精子の先体突起は卵膜を貫通し卵細胞膜に達していたが、精子と卵細胞膜との融合は起こっていないことが明らかとなった。以上の結果から、ムラサキイガイ卵の多精拒否機構は電気的な早い機構と、それに続く遅い機構とから成り、更に遅い機構は先体反応の抑制による精子結合阻止機構と卵細胞膜と精子の膜融合の阻止機構の二つから成ることが明らかとなった(図1)。 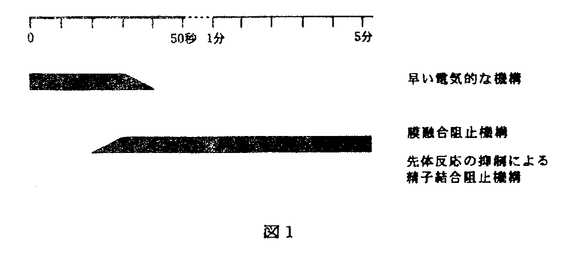 図1早い電気的な機構膜融合阻止機構光体反応の抑制による精子結合阻止機構第2部ムラサキイガイ卵における先体反応の抑制による多精拒否 図1早い電気的な機構膜融合阻止機構光体反応の抑制による精子結合阻止機構第2部ムラサキイガイ卵における先体反応の抑制による多精拒否 第2部では、ムラサキイガイ卵の遅い多精拒否機構のうち、先体反応の抑制機構について更に検討を行った。先体反応抑制の原因は、卵表の先体反応誘起に関る因子が受精後に何らかの作用を受けてその機能失うことにあると考えられる。そこで、その因子の機能を失わせる要因としてのプロテアーゼの関与を検討した。 まず、未受精卵をプロテアーゼ阻害剤存在下で受精させ、先体反応抑制機構が成立するかどうかを調べた。検討した様々なプロテアーゼ阻害剤のうち、アミノペプチダーゼ阻害剤であるamastatinとbestatinの存在下で媒精して得られた受精卵を阻害剤の非存在下で再媒精すると、先体反応の抑制に伴う精子結合の阻止が起こらず、未受精卵へ媒精した場合と同様に精子が卵に結合することが明らかとなった。同様にアミノペプチダーゼの人工基質Leu-MCAの存在下で受精させた場合も先体反応抑制機構は成立しなかった。これらのことから、この機構の成立にはアミノペプチダーゼが関与していることが示唆された。 次に未受精卵を受精させ、その懸濁液の上清のプロテアーゼ活性を調べたところ、高いアミノペプチダーゼ活性を示し、またこの活性はamastatin、bestatin、phenanthrolineなどのアミノペプチダーゼ阻害剤で阻害された。これらのことから、受精時に卵からアミノペプチダーゼ様プロテアーゼが放出されることが示唆された。 また未受精卵をこのアミノペプチダーゼを含む上清(以下fertilization product,FP)で処理したあと媒精すると、FPの濃度に依存して精子結合が阻害された。更にアミノペプチダーゼ阻害剤存在下でFPを未受精卵に作用させてから媒精すると、精子結合は阻害されなかった。同様に先体反応への影響を調べたところ、FP処理した卵に媒精した場合に先体反応率の低下が観察された。 以上の結果からムラサキイガイ卵における先体反応の抑制機構は、受精時に卵から放出されるアミノペプチダーゼ様プロテアーゼが卵表に作用することによって成立すると考えられる。 第3部ムラサキイガイ精子に存在するアミノペプチダーゼの先体反応における役割 第2部で示したように、ムラサキイガイにおいては受精後に卵から放出されるアミノペプチダーゼ様プロテアーゼが卵表に作用し、先体反応を抑制する。このことは、卵表に存在するアミノペプチダーゼ様プロテアーゼの基質が先体反応誘起に深く関っていることを示唆する。そこで第3部では、卵表のアミノペプチダーゼ様プロテアーゼの基質が先体反応誘起に関与しているかどうかを検討した。 精子の懸濁液に様々なプロテアーゼ基質を加えたところアミノペプチダーゼの基質が強く分解され、この分解活性はアミノペプチダーゼ阻害剤であるactinonin、amastatin、bestatinで抑制された。一方、精子懸濁液の上清にこの活性は無く、精子表面にアミノペプチダーゼが存在していることが示唆された。 更にアミノペプチダーゼ阻害剤存在下で媒精すると、多くの精子は先体反応を抑制されて卵への結合を阻害された。また人工基質Leu-MCAの存在下で媒精した場合も精子結合が阻害された。以上の結果から、ムラサキイガイにおいては先体反応の際に精子アミノペプチダーゼと卵表の基質との相互作用のあることが示唆された。 第2部と第3部の結果から、ムラサキイガイにおける先体反応誘起と、多精拒否機構としての先体反応抑制について、次のような仮説を提出した(図2)。卵表の先体反応誘起に関る因子は受精に伴って卵から放出されるアミノペプチダーゼの作用を受けてその機能を失う。その結果、遅れて卵表に到達した精子のアミノペプチダーゼは、卵表の基質と相互作用できず、先体反応を抑制されていると考えられ、アミノペプチダーゼの基質は先体反応誘起因子である可能性がある。 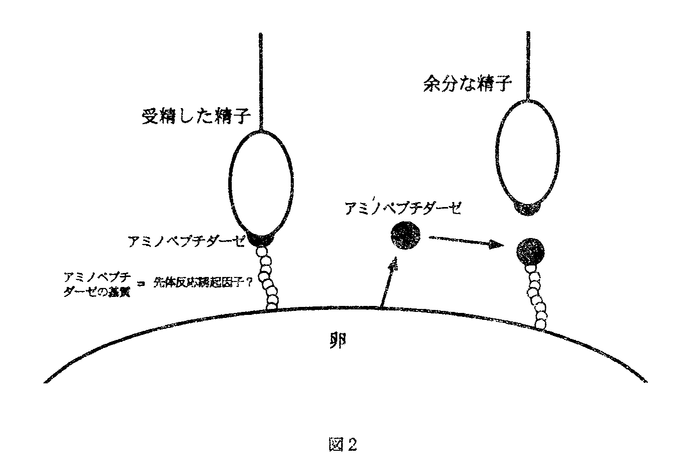 図2第4部マガキ卵における多精拒否機構 図2第4部マガキ卵における多精拒否機構 第1〜3部において明らかになったムラサキイガイ卵の多精拒否機構が、二枚貝に一般的な機構であるか否かについて知るために、マガキ卵における多精拒否機構について検討を行った。トリプシンにより未受精卵の卵膜を除去してから媒精しても多精は誘起されなかった。このことからマガキ卵では卵膜は多精拒否に積極的な役割を果たしておらず、余分な精子の排除は卵細胞膜で行われていることが示唆された。 人工海水中のNa+濃度を低くして媒精したところ多精が誘起された。このことはマガキ卵に電気的な早い多精拒否機構が存在していることを示唆する。 一方、受精卵を再び媒精すると、ムラサキイガイの場合とは異なり、再媒精した精子は受精卵への結合を阻止されなかった。次にムラサキイガイの場合と同様にFPを得て、プロテアーゼ活性を調べたところ、プロテアーゼ活性はほとんど検出されなかった。 また卵に結合した精子を電子顕微鏡で観察したところ、精子は先体反応を起こしていたが、先体突起と卵細胞膜との融合は確認できなかった。 従ってマガキ卵の多精拒否機構は(1)早い電気的な機構、(2)膜融合阻止機構の2段階から成り、ムラサキイガイでみられたプロテアーゼの作用による先体反応抑制機構を欠いていることが明らかとなった。 |