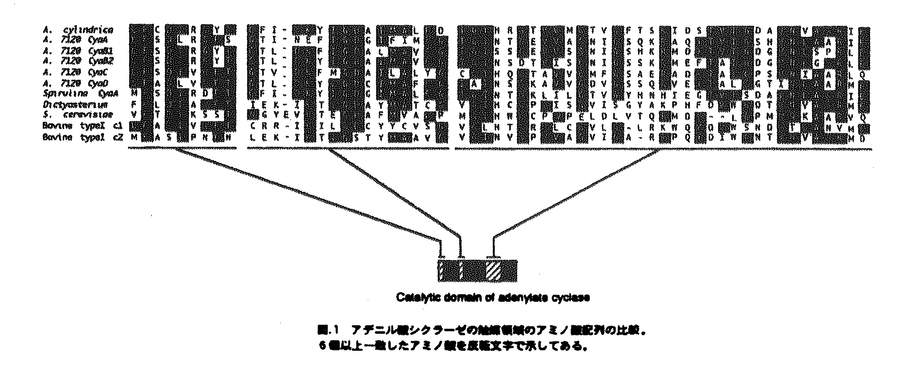
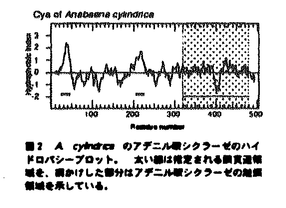
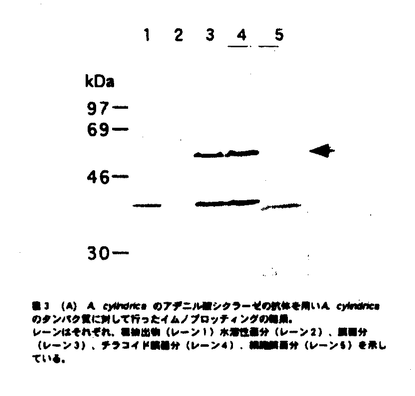
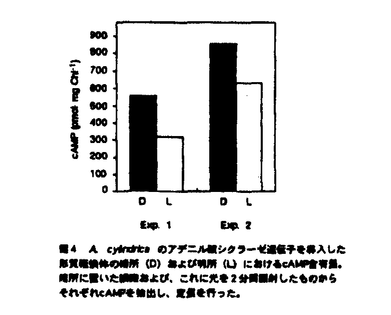
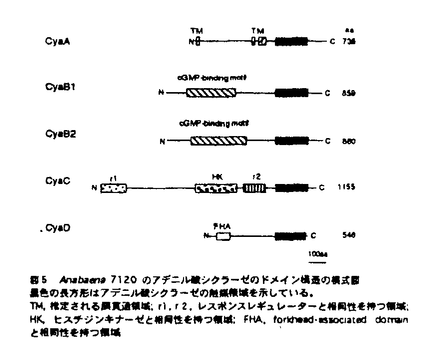
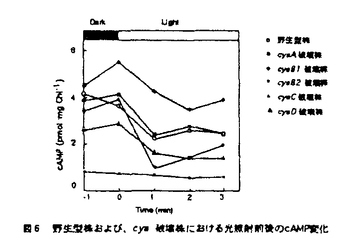
本論文は3章からなり、第1章は、Anabaena cylindricaのアデニル酸シクラーゼ遺伝子の単離、第2章は、A.cylindricaのアデニル酸シクラーゼ遺伝子のAnabaena7120における過剰発現、第3章は、Anabaena7120のアデニル酸シクラーゼ遺伝子の単離とその構造解析について述べられている。 サイクリックAMP(cAMP)は細胞内情報伝達物質として良く知られており、糸状性ラン藻Anabaena cylindricaにおいては、cAMPが光やpHなどの環境条件の変化にともなって変動することが報告されている。cAMPの機能を解析するためにはcAMPの合成酵素であるアデニル酸シクラーゼについての情報が必要とされる。これまで、ラン藻のアデニル酸シクラーゼの遺伝子は単離されておらずその構造は不明であった。本研究では二種のAnabaenaよりアデニル酸シクラーゼ遺伝子を単離し、その構造の解析を行った。 第1章を要約は以下のようである。大腸菌のアデニル酸シクラーゼ遺伝子欠損株のラクトース代謝能の相補を指標にしてラン藻A.cylindricaのゲノムDNAライブラリーをスクリーニングし、アデニル酸シクラーゼ遺伝子を含むDNA断片を一つ取得した。A.cylindricaのアデニル酸シクラーゼは動物や酵母などの真核生物のアデニル酸シクラーゼの触媒領域と高い相同性を示した。一方、A.cylindricaのアデニル酸シクラーゼは大腸菌のアデニル酸シクラーゼとは相同性を示さなかった。このアデニル酸シクラーゼはN末端とタンパク質の中央付近にそれぞれ膜貫通領域と思われる疎水性の領域を含み、C末端側にアデニル酸シクラーゼの触媒領域を持つ、全体として膜結合型の構造を持つことが明らかとなった。また、このアデニル酸シクラーゼは、チラコイド膜画分に局在していることが明らかとなった。 第2章の要約は以下のようである。宿主ベクター系が確立されているラン藻Anabaena sp.strain PCC7120(Anabaena7120)を用いてA.cylindricaのアデニル酸シクラーゼの過剰発現を試みた。作成した形質転換体は野性型のAnabaena7120の約170倍のcAMPを蓄積しており、さらに暗所に置いた形質転換体の細胞に光を照射するとcAMPレベルが減少した。イムノブロッティングの結果、導入したアデニル酸シクラーゼは細胞膜とチラコイド膜の両方に存在していた。また、この形質転換体では糸状体が切れて細胞がばらばらとなりこれらが接着した集団を形成することが観察された。 第3章の要約は以下のようである。遺伝子導入の容易なラン藻Anabaena7120からアデニル酸シクラーゼ遺伝子を単離した。その結果、5種類のDNA断片が単離された。塩基配列を調べたところ、これら5種類のDNA断片上にはそれぞれ既知のアデニル酸シクラーゼと高い相同性を持つタンパク質をコードするオープンリーディングフレームが存在することが明らかとなった。よって、これらをアデニル酸シクラーゼ遺伝子と同定し、それぞれcyaA,cyaB1,cyaB2,cyaC,cyaDと名付けた。 cyaAは735アミノ酸残基からなるタンパク質をコードしていた。CyaAはC末端側に触媒領域を持ち、N末端とタンパク質の中央付近に2箇所に分散した疎水性領域を持つ膜蛋白質であると想像される。cyaB1とcyaB2はそれぞれ859および860アミノ酸残基からなる親水性のタンパク質をコードしていた。CyaB1とCyaB2は互いに全長にわたって相同性を示した。cyaCは1155アミノ酸残基からなる親水性のタンパク質をコードしていた。CyaCはN末端から順にバクテリアの二成分制御系のレスポンスレギュレーター、ヒスチジンキナーゼタンパク質、レスポンスレギュレーターと相同性のある配列を持ち、C末端側にアデニル酸シクラーゼの触媒領域を持っていた。cyaDは546アミノ酸残基からなるタンパク質をコードしていた。CyaDのN末端には、ある種の転写調節因子およびキナーゼに存在しているForkhead-associated domainと呼ばれる領域と相同性を示す配列が存在していた。通常の培養条件において全てのアデニル酸シクラーゼ遺伝子が発現していることがRT-PCR反応により明らかとなった。また、いくつかのアデニル酸シクラーゼ遺伝子の破壊株において細胞のサイズに野性型株との間で違いが見られた。 以上、本研究において、初めてラン藻のアデニル酸シクラーゼ遺伝子の構造が明らかとなった。さらにラン藻は複数のアデニル酸シクラーゼ遺伝子を持つことが明らかとなった。これらの遺伝子は互いに異なった構造と発現パターンを持つことから、異なる情報伝達系に関与している可能性が示唆された。このように本論文はこれまで知られていなかった、ラン藻におけるアデニル酸シクラーゼ遺伝子に関して新しい知見を与えるものであり、科学的に高い価値がある。 よって、博士(理学)の学位を授与できると認める。 |