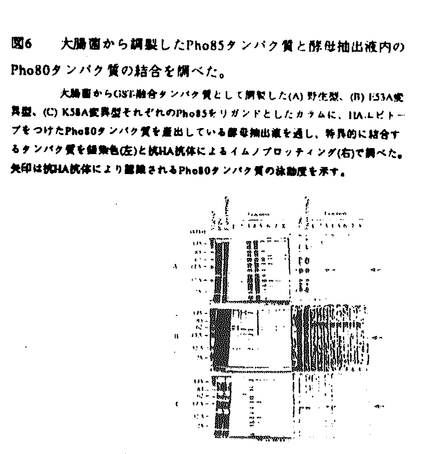
本研究は、出牙酵母のサイクリン依存性プロテインキナーゼのひとつであるPho85キナーゼによる多面的な形質支配の機構を明らかにすることを目的として行われた。第一章では、遺伝学的方法によりpho85変異が多面的な形質変化を引き起こす現象を明らかにし、第二章では、Pho85蛋白質の機能ドメインを解析し、第三章では、Pho85キナーゼによる細胞周期制御について論じられている。詳細は以下のとおりである。 第一章Pho85キナーゼによる多面形質支配 pho85遺伝子破壊株では、Pho5酸性フォスファターゼの活性が培地内の無機リン酸イオン濃度に依存せず構成的になる。このPhO5遺伝子の構成的発現以外に、pho85遺伝子破壊株は、増殖の遅延、ガラクトースや非発酵性の炭素源の資化異常といった表現型を示した。これらpho85遺伝子破壊株の表現型は全て、pho4遺伝子破壊により抑圧された。PHO4遺伝子はPHO5遺伝子の転写活性化因子であり、Pho85キナーゼによるリン酸化を受け不活化することが明らかになっている。つまり、Pho85キナーゼがPHO系のみならず、正常な増殖や炭素源の資化をもPho4タンパク質を介して調節している可能性を示唆する。Pho80タンパク質はPho85キナーゼのサイクリンとして機能する。pho80遺伝子破壊株はpho85遺伝子破壊株と同様にPho5酸性フォスファターゼの活性が構成的になるが、非醗酵性の炭素源の培地上では生育可能であった。このことからPho85キナーゼがPho80タンパク質以外のサイクリンと結合して炭素源の資化を制御し、しかも炭素源の資化の情報もまたPho4タンパク質を介している可能性が考えられる。 第二章Pho85キナーゼの機能ドメイン ATP結合領域、PSTAIRE配列、活性化リン酸化部位などCdkの機能に重要と考えられているアミノ酸配列は、Pho85キナーゼにおいても保存されている。これらのアミノ酸残基に点変異を導入して変異Pho85キナーゼを作成し、in vivoではPho5酸性フォスファターゼの活性と非醗酵性の炭素源上での生育状況、in vitroではキナーゼ活性を指標にPho85キナーゼの機能ドメインについて調べた。ATP結合領域内のY18,22F変異、PSTAIRE配列内のE53A変異、及びK58A変異はin vivoでPho85キナーゼとして機能しなかった。これらの変異はin vitroでのキナーゼ活性も低下していた。PHO80遺伝子の発現量を増やすことで、Y18,22F、K58A変異の活性はin vivo及びin vitroで回復したが、キナーゼ活性に必要な残基であるE53A変異の活性は失われたままであった。Y18,22F変異とK58A変異による活性の低下は、Pho80タンパク質との結合が弱まってサイクリンによる活性化を受けにくいためと考えられる。活性化リン酸化部位に相当するF165S166S167への変異はPho85キナーゼのin vivoでの機能に影響を及ぼさず、in vitroでも野生型と同程度のキナーゼ活性を持っていた。他のCdkのようなCdk Activating Kinase(CAK)のリン酸化による活性化の機構がPho85キナーゼに存在するかどうかはまだ明かではないが、S166S167残基はPho85キナーゼの機能に重要ではないことが言える。 Y18,22F変異とK58A変異の活性の低下がPho80タンパク質との結合が弱くなっているためと予想し、変異Pho85キナーゼとPho80タンパク質との物理的な相互作用について調べた。大腸菌から調製した野生型、E53A変異、K58A変異のそれぞれのPho85タンパク質をリガンドとしたカラムに、Pho80タンパク質を過剰生産している酵母抽出液を通したところ、野生型及びK58A変異Pho85タンパク質のカラムでは特異的なPho80タンパク質の結合が観察され、E53A変異ではPho80タンパク質の結合は見られなかった。また、Pho80タンパク質が特異的に溶出される画分にはPho80タンパク質以外のタンパク質の存在が観察された。このことはPho85キナーゼがPho80タンパク質を含むいくつかのタンパク質と結合して複合体を形成して機能していることを示唆する。そこで、酵母細胞内で機能しているPho85キナーゼが含まれるタンパク質複合体を解析した。グルタチオン-S-トランスフェレース(GST)融合タンパク質としてPho85キナーゼを産出している酵母細胞抽出液を用い、抗GST抗体をリガンドとしたカラムに特異的に結合する免疫タンパク質複合体を調べた。野生型Pho85キナーゼの含まれる画分にはそれ以外のタンパク質も特異的に溶出されてきたが、機能を持たないE53A変異の場合あるいはpho80変異株から調製した細胞抽出液の場合には、野生型のGST-Pho85キナーゼの含まれる画分に含まれていた他のタンパク質は観察されなかった。つまり、酵母細胞内でPho85キナーゼが機能するためには、Pho80タンパク質を含んだタンパク質複合体形成が必要であり、Pho85キナーゼはPho80タンパク質を介してタンパク質複合体と結合することが予想される。 第三章Pho85キナーゼによる細胞周期制御 Pho85キナーゼが細胞周期の進行に時期特異的に作用しているのであれば、細胞周期阻害剤に対する感受性が変化すると考え、細胞周期阻害剤のpho85遺伝子破壊株への影響を調べた。すると、pho85遺伝子破壊株はヒドロキシウレア(HU)に対して野生型より高い感受性を示した。pho80遺伝子破壊株は野生型株と同程度の、またpho85pho4二重遺伝子破壊株はpho85遺伝子破壊株と同程度の感受性を示したことから、HU存在下ではPho85キナーゼはPho80タンパク質以外のサイクリンによる活性化を受け機能することが予想される。紫外線照射やメチルメタンスルホン酸処理に対するpho85遺伝子破壊株の感受性が大きく変化しないことや、HU処理をしてもpho85遺伝子破壊株の生存率は顕著には低下しないことから、pho85遺伝子破壊株ではチェックポイントコントロール機構は機能しており細胞周期停止からの回復過程に障害があることが考えられる。 HUはDNA合成を阻害して細胞周期をS期で止めることが知られているが、DNA合成はCdc28-Clb5,6複合体が促進し、このCdc28-Clb5,6複合体の活性はSic1タンパク質により阻害される。HUに対する感受性が高くなった原因として、pho85遺伝子破壊株ではSic1タンパク質が適切に分解されずにCdc28-Clb5,b複合体の活性が充分に上昇しないことを考え、Sic1タンパク質のpho85遺伝子破壊株内での安定性を調べた。すると、野生型株に比べpho85遺伝子破壊株ではSic1タンパク質はより長時間存在した。Sic1タンパク質は、リン酸化の修飾が引金となってユビキチン化され速やかに分解される。pho85遺伝子破壊株でSic1タンパク質がより安定に存在する可能性として、Pho85キナーゼによるリン酸化がユビキチン化を介した分解に関係していることを予想し、Sic1タンパク質がin vitroでPho85キナーゼの基質となるかを調べた。大腸菌から調製したSic1タンパク質は、Pho85キナーゼによりリン酸化された。pho85遺伝子破壊株内でのSic1タンパク質の安定化とin vitroでのPho85キナーゼによるリン酸化より、CKIの分解を制御するCdkとしてPho85キナーゼが細胞周期を調節する可能性が示唆される。 以上のように、本論文はPho85キナーゼの新規な細胞機能を明らかにしている。とくに、細胞周期制御について新しい作用機構をs提案している。発表論文は共著であるが、実験計画とその遂行は申請者によって行われたものである。このような評価に基づき、本研究は博士(理学)の学位に十分値するものであることが審査員全員の一致により認められた。 |