軟体動物は外骨格として硬組織(貝殻)を持つため化石記録が極めて豊富な動物群である。特に、腹足綱の中でも原始腹足類(Archaeogastropoda)はカンブリア紀後期より現世まで連続的に化石記録を持つ主要な分類群の一つであり、約80科(現生では約40科)が含まれる。この類は解剖学的には多くの原始的形質を持つことが知られており、例えば外套腔や内部器官の対称性は軟体動物の共通祖先より保存されている原始性の反映であると考えられてきた。よって原始腹足類は腹足類全体の起源を探る上でも、さらにはより派生的な特徴を持つ腹足類の進化を考える上でも系統学的に重要な位置を占めている。 現生腹足類の高次分類群の体系は主に軟体部の特徴に基づいて議論されてきた。近年の系統分類においては解剖学の進歩により多くの新しい成果が発表されつつあるが、依然として多くの分類群において詳細な解剖データが不足しており高精度の系統構築上の障害となっていた。よって本研究では幅広い分類群にわたって新たに高精度の解剖学的データを収集し、それらを詳細に比較分析することにより原始腹足類の系統関係を明らかにし、殻体形質も含めた形態進化についての仮説を提唱することを目的とした。 具体的な手順としては、まず比較解剖の結果から分岐分析に基づき系統関係を推定し、形質変化の極性についても考察した後、軟体形質から得られた分岐図上に殻体形質(原殻・殻体構造)の分布を重ね合わせることにより系統的対応関係を検証した。解剖は実体顕微鏡下における肉眼解剖、パラフィン包埋連続切片、SEMによる微細構造の観察を総合して行い、17科23属25種を対象に、一種につき200以上の形質に関する解剖学的データを得た。次に系統解析では、離散的形質状態を示す形質について、微細構造と位置の基準(神経支配の器官特異性など)に基づき相同性を慎重に吟味し、系統解析に利用可能な形質として93形質を抽出した。それらの内約20形質が過去の系統的研究では検討されたことのない新形質であった。分岐分析は系統解析ソフトPAUP(=Phylogenetic Analysis Using Parsimony)ver.3.1を用い、外群としてヒザラガイ・ネオピリナ・オウムガイを含めて28分類群・93形質について発見探索法(heuristic search)により最短系統樹を計算した。 分岐分析の結果、以下のような系統関係が明らかになった(図1)。(1)Patellogastropodaは外群と複数の原始形質を共有しており、腹足類の中では最初に分岐する。特に、歯舌や顎板を含む消化系がこの関係を支持する形質として重要である。(2)Patellogastropoda以外の腹足類が単系統群を形成する。これはPonder and Lindberg(1996)のOrthogastropodaに対応する。(3)Vetigastropoda(s.s.)(=Zeugobrachia+Trochoidea)は単系統である。さらに、LepetodriloideaとSeguenzioideaがより大きな単系統群を形成し、これらはVetigasatropoda(s.l.)としてまとめることができる。(4)Cocculinidaeの系統上の位置については2通りの可能性があり、今回の分析結果からはそのどちらが妥当であるかは特定できなかった。よって厳密合意樹ではOrthogastropodaの関係は完全に多分岐となった。(5)Neritimorpha(陸棲種を含む)の単系統性は多くの形質によって支持される。特に、口球、消化管、生殖器系の形質が重要である。(6)ArchitaenioglossaはNeotaenioglossaと複数の派生形質を共有しており、Caenogastropodaとしてまとめられる。従って、Architaenioglossaは原始腹足類に含めることは出来ない。しかし、神経系においてはNeritimorphaと共有される形質があり、よってCaenogastropodaの中でも原始的な位置を占める。(7)以上の結果より、Thiele(1925)の提唱したArchaeogastropodaは単系統ではなく側系統であることが明らかになった。従って、Archaeogastropodaは分岐群(clade)ではなく、梁舌型または扇舌型の歯舌を持つ段階群(grade)として定義される。 次に各形質ごとに分岐樹上での形質状態の変化の極性を追跡した結果、軟体部の進化に関して次のような傾向が明らかになった。(1)Patellogastropodaが真に最も原始的な腹足類であるならば、Zeugobranchiaに見られる外套腔の対称性は2次的であると解釈される。この考えは従来の説とは全く異なるが、個体発生の基準からは支持される。よって腹足類の祖先は既に非対称な外套腔と内部器官を持っていたと考えられる。(2)多板類・単板類と共通性の高い原始的な口球筋肉が原始腹足類ではよく保存されていることが明らかになった。一方、筋肉系の著しい特殊化・単純化がCaenogastropodaの共通祖先で起きたものと思われる。口球軟骨・消化管においても同様に単純化の傾向が明らかである。(3)心臓や体腔器官の非対称性は完全にモザイク状の形質分布を示す。よって非対称性(右側の退化)は複数の分類群において独立に進化したものと思われる。 次に、軟体部から推定される分岐関係を殻体形質の類型と対照し、殻体形質の高次分類上の評価を試みた。その結果、原殻形質は外形により次の4類型に分けられ、それらは解剖学的に定義される高次分類群と基本的によく対応することが明らかになった。(a)壺型:前後に長く左右対称。Patellogastropodaのみに見られる。内臓塊先端部が吸収された後に隔壁が形成され、殻長1〜2mmの段階で胎殻は脱落する。(b)少旋型:胎殻は形成初期から巻き始め、非対称になるものが多い。Cocculiniformia,Vetigastropoda等(すなわちNeritimorpha以外のRhipidoglossa)に見られる(例外的にMacroshismaスカシガイは左右対称であり全く巻かない)。(c)多旋型:捕食型幼生期に原殻IIが形成され、付加成長による成長線を残す。水棲アマオブネガイ目(Neritimorpha)に見られ、深海〜淡水に棲息する種(複数科)間で原殻形態は酷似する。内部では殻軸の再吸収が起こる。(d)陸生型:陸棲アマオブネガイ目は卵殻内で発生するため外部形態は著しく特殊化するが、内部は殻軸が再吸収される点で(c)と共通である。 また、以上の結果から原殻形質の形態進化に関して以下のような結論が得られる。(1)軟体部から推定される進化過程との対応から、原始腹足類の原殻は、左右対称型→少旋型→多旋型という過程を経て進化したと解釈される。(2)Neritimorpha以外の原始腹足類では原殻表面は稜柱構造からなる不規則な彫刻に覆われ、付加成長による成長線を欠く。ProtoconchIIの形成はアマオブネガイ目より派生的な腹足類のみにみられる形質である。(3)壺型のPatellogastropodaには全個体発生過程を通じて全く左右対称な種があり、幼生期に起こる軟体部の捻れ(Torsion)と貝殻の巻きが独立に生じていることを示す。(4)上記の原殻の類型化は収斂によって生じた笠型貝類の同定に極めて有効である。特に、殻頂が前寄りに位置するPatellogastropodaは、後ろ寄りの他の腹足類(Orthogastropoda)とは明確に区別される。 一方、殻体構造においても特定の構築構造の組み合わせが比較的低次(科より下のレベル)の高次分類群に見られる。腹足類の殻体形質に関して最も興味深い問題は柱状真珠構造が原始性を反映するかどうかという点である。真珠構造は腹足類の中ではVetigastropodaのみにしか見られない特異的な形質であるが、外群も含めた分岐図上ではVetigastropodaと頭足類の間で同形形質(homoplasy)になる。しかし、構造的さらには個体発生的には両者の(共有原始形質としての)相同性は否定できない。さらに、Vetigasatropodaの内部にも真珠構造を欠く分類群が散見される。これらのことから真珠層の欠如は独立に複数回進化したものと解釈され、多くの場合は小型化による二次的な変化であるとみなされる。殻体構造の形質は軟体部に基づく高次分類群の関係とは整合性が低く、これは恐らく殻体形質の著しいモザイク状進化に起因するものと思われる。しかし、低次の分類群では形態型は同定に利用可能であり、原殻形質・筋肉痕・終殻に見られる形質とともに分類形質としての価値は高い。 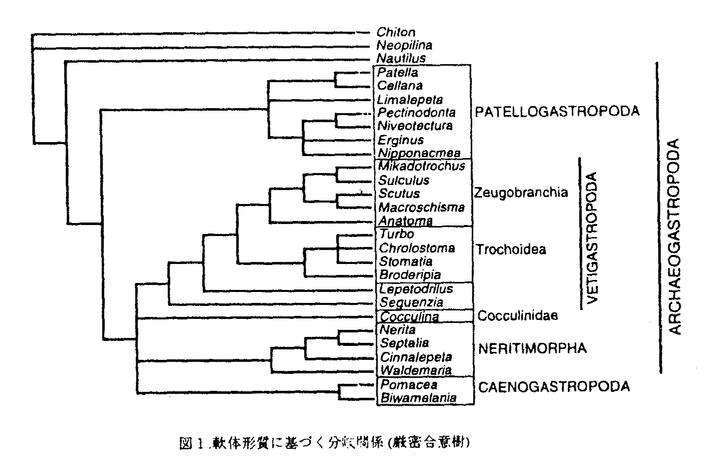 図1.軟体形質に基づく分岐関係(厳密合意樹) 図1.軟体形質に基づく分岐関係(厳密合意樹) |