従属栄養性微小鞭毛虫類(HNF)はAzamらが提唱した微生物環説によって海洋生態系での存在が重要視されるようになり、細菌捕食者として注目された。現在までに生態系におけるエネルギー流量の見積りに関する研究が行われているが、捕食形態、分裂様式などの個体レベルの知見が極めて少ない。海洋生態系の物質輸送を明らかにする上では重要な生物である。 本論文は、海洋におけるHNFの分布密度、個体レベルにおけるHNFの特性として捕食様式と分裂様式を解析することによりHNFの生物学的な知見について言及したもので次の5章からなる。 第1章は序章で、HNFは分類学的に多岐に位置付けられ、生態学的知見は多く報告されているが、生物学的知見について極めて少ないことを述べている。 第2章はHNFの単離と固定条件の検討である。採集は東京湾湾奥部から行い、採取できた2〜3種の鞭毛虫で固定条件の検討に使用している。さらに段階希釈法を用いて単離できたActinomonas sp.(体長約7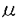 m、鞭毛が1本)とBodo sp.(体長約5m、鞭毛が2本)の2種である。固定条件の検討では(1)固定した標本を染色しないもの、(2)固定により可能な限り細胞消失の起こりにくいもの、(3)実験終了後の廃液処理が簡便なものとして、グルタルアルデヒドとホルムアルデヒドを選択していた。固定直前に対する固定後の細胞密度の細胞消失率により比較検討していた。両固定液による鞭毛虫の形状比較などから、鞭毛虫密度を測定の最適固定濃度はグルタルアルデヒドで最終濃度1%固定であることを明らかとしている。 m、鞭毛が1本)とBodo sp.(体長約5m、鞭毛が2本)の2種である。固定条件の検討では(1)固定した標本を染色しないもの、(2)固定により可能な限り細胞消失の起こりにくいもの、(3)実験終了後の廃液処理が簡便なものとして、グルタルアルデヒドとホルムアルデヒドを選択していた。固定直前に対する固定後の細胞密度の細胞消失率により比較検討していた。両固定液による鞭毛虫の形状比較などから、鞭毛虫密度を測定の最適固定濃度はグルタルアルデヒドで最終濃度1%固定であることを明らかとしている。 第3章はHNFの海洋における分布様式の調査である。鹿島灘10測点、太平洋の亜寒帯外洋域1測点、亜熱帯中央水域3測点と赤道湧昇域2測点で微小プランクトンの鉛直分布を調査していた。鹿島灘の調査では、HNF、ANF(独立栄養性微小鞭毛虫類)、ラン藻類および細菌ともに50m以浅に多く存在したが、50m以深では減少する傾向が見られた。HNFはANFより上側に偏って存在する傾向が認められた。これら微小プランクトン群の各測点間の水柱積算細胞数は水塊によって大きく変化していなかった。太平洋の亜寒帯外洋域、亜熱帯中央水域および赤道湧昇域の調査では、鹿島灘と同様に水温躍層以深より各微小生物の密度は減少していた。HNFはANFより浅い層に偏って分布し、ラン藻類の極大層は亜表層クロロフィル極大層とほとんど一致していた。海域によるHNFの分布には差異が見られないことを明らかにしている。 第4章は高速ビデオカメラを用いた捕食様式の観察である。単離したActinomonas sp.とBodo sp.について水温および塩分条件を変化させ、培地中に存在する細菌を捕食させていた。捕食様式はそれぞれ4段階に区分できることを明らかにしている。Bodo sp.捕食には、’包み込み’と’吸い込み’の2様式があり、吸い込みは包み込み捕食時間に比べ有意に短かく、水温上昇とともに長くなっていた。また捕食中に接近する他の細菌には反応を示していなかった。一方Actinomonas sp.の捕食には、’包み込み’様式のみ観察され、捕食中接近する細菌を連続的に捕獲し、次の捕食まで細菌を触手に保持していた。捕食時間は水温、塩分変化に対し有意差が認められず、Bodo sp.に比べ約25倍も長かったが、この原因として両種で食飽形成の細胞膜再生速度に差があったためと考察していた。半数が10分間に捕食せず、捕食間隔には周期性が認められなかった。接近する細菌の頻度には個体によるばらついていたことを明らかにしている。 第5章は分裂様式の観察である。単離したActinomonas sp.とBodo sp.の2分裂を観察していた。Actinomonas sp.ついては、7段階に区分でき、分裂時間が10〜20分であることを明らかにしている。一方、Bodo sp.については、6段階に区分でき、分裂時間が鞭毛が停止してから5〜10分であることを明らかにしている。Bodo sp.の分裂には出芽形式の分裂があることを明らかにしている。さらに、両属とも体型を変化させており、Actinomonas sp.については4型式、Bodo sp.については8型式あることを明らかにしている。 以上要するに、本論文は海洋生態系の微生物環における主要な微小生物でありながら、生物学的知見の少なかった従属栄養性微小鞭毛虫類の個体レベルでの特性に着目し、分布様式、捕食様式および分裂様式などの特性について新知見を得たもので、学術上、応用上寄与するところが少なくない。よって審査委員一同は本論文が博士(農学)の学位論文として価値あるものと認める。 |