Brachionus属の海産ワムシは海産魚類及び甲殻類の人工種苗生産用初期餌料として最も重要な動物プランクトンであり、全国各地の種苗生産機関では、ほぼ30年にわたって大量培養が行われてきた。しかし、ワムシの培養においては未だ原因不明とされる急激な個体密度の低下や大量死亡現象がしばしばみられ、計画的種苗生産にとって大きな支障となっていることも事実である。 本研究では、従来の水質化学的研究とは異る視点に立ち、ワムシ飼育水中に構成される微小生物生態系を環境の「演出者」としてとらえ、そのうちでも大きなバイオマスを占めると考えられる原生動物、とくに繊毛虫の機能をワムシ培養の成否との関わりにおいて論じた。すなわち、第1章では給餌率を変えたワムシ培養槽において原生動物相を調査し、第2章では、最も大量に出現する繊毛虫Euplotes vannusを分離培養ののち、ワムシ培養系に添加した場合のワムシ増殖曲線の変化からE.vannusがワムシに与える影響を明らかにした。 また、その影響を与えるメカニズムについては、第3章で培養槽内生態系に存在する様々な有機物に対するワムシ及びE.vannusの利用形態の違いを明らかにすることによって考察した。さらに第4章では、新たに発見した,ワムシを死に至らしめる原生動物について生物学的特性を調べワムシ大量培養への阻害機構を考察した。 1.ワムシの培養における原生動物相の遷移 実際のワムシ大量培養に準じ、藻類を培養水とともに水槽に導入し,ワムシ20個体/mlを接種した。その後藻類が食べ尽くされて以降の餌料を、一般的な餌料であるパン酵母に切り換えた。 ワムシの密度が100個体/mlに到達するまでの培養初期には主に鞭毛虫が出現し、それ以後の,対数増殖期と安定期を含めた培養中期には繊毛虫が優占したが、培養後期には鞭毛虫も再び出現した。繊毛虫の種類はUronemaとEuplotesであった。パン酵母の給餌量毎に見ると、水量20Lに毎日11.2g(107cells/mlに相当)を投入した1/1区では、ワムシ密度は4日目から100個体/mlを超えて1000個体/ml前後に達したが、この状態の維持は20日間に及んだケースと、わずか8日間ののち急激に落ちるケースに分れた。原生動物の出現量は培養後期に増えた。酵母濃度2/1区では、ワムシは順調に1000個体/mlを超えたものの、15日目頃から急激に落ちる場合があった。酵母濃度1/2区では、ワムシ密度は数100から1000個体/mlまでを比較的安定して維持した。 原生動物として卓越した繊毛虫Uronema及びEuplotesについては競合関係が見られることなどからniche(生態学的地位)は等しいものと考えられたが、どちらか先に約1000cells/mlを超えた方が優占種になることが多かった。細菌数及び繊毛虫量は酵母投与量が多いと高くなった。餌料競合関係ではワムシが繊毛虫より強いと考えられたが、2/1給餌区で培養後期にみられたように環境悪化によってワムシが死亡し、原生動物のみが卓越することもあった。 2.繊毛虫,Euplotes vannusのワムシの増殖に対する影響 生態学的地位が等しいと考えられた繊毛虫Euplotes(長径約75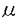 m)とUronema(長径約20m)のうち、より大きいE.vannusを代表種としてワムシ培養水槽から分離・培養したのちワムシの培養に加え、その影響を調べた。 m)とUronema(長径約20m)のうち、より大きいE.vannusを代表種としてワムシ培養水槽から分離・培養したのちワムシの培養に加え、その影響を調べた。 2種の藻類NannochloropsisまたはTetraselmisを給餌したワムシ培養に対し、その「誘導期」、「対数増殖期」及び「定常期」に1000cells/mlのE.vannusを加えたとき、Nannochloropsisを用いたワムシ培養では、添加時期にかかわりなくE.vannusは接種密度を大きくは超えない、あるいは1/2程度にまで減少したが、対数増殖期に添加された2例のうち,E.vannusが2000cells/ml程度まで比較的早く到達した場合には、ワムシの増殖を抑制し、また、定常期に添加された場合にはワムシ密度を低下させた。 Tetraselmis給餌によるワムシ培養では、E.vannusの到達密度は1mlあたり数千から1万個体近くにもなり、特にワムシの誘導期に添加された場合は2-3日後には2000cells/mlを超え、ワムシ増殖を抑制した。ワムシの対数増殖期あるいは定常期に添加された場合、E.vannusははじめ増殖しないが、やがてワムシ培養の経過とともに個体密度を増加させ、1000cells/mlを超えて急激な増殖に転ずるとワムシ密度を減少させた。 これらの結果から、E.vannusは1000cells/mlを超えて増殖傾向にあるときワムシの増殖を抑制するが、自身の増殖にとってNannochloropsisは餌料として好適ではなく、一方Tetraselmisは好餌料ではあるが、ワムシの増殖が盛んな場合には先にワムシに摂餌されてしまうものと考えられた。 3.ワムシと繊毛虫における有機物利用特性と増殖の機構 本章では、前章までの実験によって示唆された,ワムシとE.vannusの間に見られる餌としての有機物利用をめぐる競合関係について、そのメカニズムを知ることを目的とした。ワムシ培養槽内に存在する有機物を溶存態有機物(DOM),生きた餌料(live-Particulated Organic Matter,l-POM),変性餌料(死亡によるもの…d-POM及び細菌が付着し分解過程にあるもの…b-POM),ワムシ糞(f-POM)に分けて定義し、人工的に調製したこれらをワムシおよびE.vannusに与え、それぞれの利用効率を評価した。なお、各々のPOMの利用は、安定同位体15Nをトレーサーとして定量化した。 その結果、E.vannusはDOMを利用できないと考えられ、また一般的なワムシ用餌料であるNannochloropsis,Tetraselmis,パン酵母のいずれについても生きた新鮮な細胞(l-POM)に対する濾水率は検出できないか、またはきわめて低かった。 この結果は増殖率による評価とも一致し、Nannochloropsisのl-POMでは個体密度は増加せず、Tetraselmisの場合でも藻の活性が低下したと思われる5日目以降にE.vannusは増殖を始めた。これに対してd-POM及びb-POMでは、E.vannusは接種直後から急激に個体数を増加させた。 15Nをトレーサーとした実験からは,E.vannusがd-POMを利用する速度(%body N/h)はNannochloropsisとTetraselmisに対して0.91及び0.94にすぎないが、ワムシ糞からは10.39及び1.50を取り込むことが明らかになった。しかし、抗生物質存在下で採取されたワムシ糞からは2.25及び1.06しか取り込まず、E.vannusの増殖が主に細菌食によっていることか明らかになった。 これら有機物利用の特性からワムシ培養槽中で繊毛虫が増殖する機構は次の様に解析された。(1)誘導期にはワムシによる摂餌圧が低く、残餌になって活力低下あるいは死亡した藻類が増加し、これらが繊毛虫の餌料となる。また、これらに細菌が付着・増殖することによってさらに餌料価値が高くなる。(2)対数増殖期はワムシによる摂餌が盛んであり、通常残餌は発生しないが、細菌の付着基質として優れているワムシ糞の発生は多い。しかし、これの捕食についてはワムシの細菌食性と競合する。(3)定常期には槽内の有機物蓄積が多く,細菌の生産が盛んであり、本来細菌食性と考えられる繊毛虫の増殖が促進される。 また、繊毛虫がワムシ増殖を阻害する機構については、細菌食に関する競合、溶存酸素消費と貧酸素耐性の違いなどが、ワムシの生理的な状況と相まって作用することと考えられた。 4.餌料培養槽より分離した太陽虫Oxnerella maritimaのワムシに対する致死作用 日本栽培漁業協会玉野事業場でワムシの拡大培養不調時に分離された直径10-15mの太陽虫はワムシ飼育後の海水を培地とし,餌料としてChlamydomonasを与えるか、またはイースト添加によって細菌の生産が促進される場合,106cells/ml程度まで増殖したが,NannochloropsisあるいはTetraselmis給餌下では増殖しなかった。この太陽虫が共存する場合,ワムシは遊泳力を失って沈降,死亡した。また,24時間後のLC50は104cells/mlであった。維持培養から大量培養へ移行する際,通常は餌料をNannochloropsisからパン酵母に切り換えるが,このことが細菌食性をもつ本種の増殖を促し、培養の不調を招いたものと考えられた。本種は微細構造などからOxnerella maritimaと同定されたが、海産ワムシに対する致死性原生動物としての初記載である。 また、致死作用の機構については、太陽虫のメタノール抽出物のSephadexによる特定の画分に致死因子が存在した。 繊毛虫の増殖による、ワムシ培養の不調ならびに太陽虫による急激なワムシの斃死が、いずれもそれら原生動物の細菌食性と深く関っていることから考えて、実際の大量培養では、適正な給餌、定期的収穫による飼育水の抜き取りを基本とする有機物負荷のコントロールが重要と考えられる。また、繊毛虫とワムシの生態学的地位が等しいことから考えると、ワムシの摂餌活性低下を招き易い水温・塩分変化を最小限に抑えることにより、微生物生態系内でのワムシへの有機物フローを確保・維持することも重要と考えられた。 |