下垂体前葉は産生するホルモンにより、少なくとも5つの細胞種に分類される。GH細胞、PRL細胞、TSH細胞、ACTH細胞、LH/FSH細胞である。下垂体前葉は胎生期に外胚葉由来の原基ラトケ嚢から発生する。それぞれのホルモン分泌細胞の分化や増殖は、近接する細胞同士の相互作用や視床下部由来因子の暴露による複雑なカスケードにより調節されると考えられる。 細胞表面には大別して2種類の分子が発現している。1つは全ての細胞に共通して発現する分子であり、もう1つは特殊な細胞のみに発現されその細胞の特徴的な機能を反映する分子(分化抗原)である。神経細胞やリンパ球細胞における分化抗原の探索及びその機能の解析により、神経系や免疫系の複雑なメカニズムが解明される一方、内分泌組織における分化抗原の解析は十分ではない。従来、下垂体前葉細胞の分化はホルモン分泌の開始点を指標に研究されてきた。本研究は、下垂体前葉GH/PRL細胞系列に特異的に発現する細胞表面分子(糖タンパク質・糖脂質)に着目し以下の研究を行った。 GH3細胞はGHとPRLの両ホルモンの分泌能を有するラット由来の腫瘍細胞である。TRH、サブスタンスP、アンギオテンシンII、インターロイキン6、EGFなど、正常下垂体細胞GH/PRL細胞系列が反応する因子に対する反応性も保持しているため、GH/PRL細胞系列の分泌機構や分化機構の研究の格好なモデル系として利用されている。 第1章では、トリプシン感受性細胞膜蛋白質(TSMPs)がPRL細胞とGH細胞の高い基礎分泌能に関わっている可能性を示し、基礎分泌能に関わるGH/PRL細胞系列特異的な細胞表面膜蛋白質の存在を提唱した。第2章では、GH/PRL細胞系列の分化抗原としてのTSMPsを探索するアプローチの一つとして、GH3細胞を抗原として単クローン抗体を作製し、その性状解析を行った。第3章では、分化抗原の候補として挙げられる酸性糖脂質(ガングリオシド)について、GH3細胞における構成成分を解析し、PRL細胞様細胞への分化との関わりを考察した。 第1章トリプシン前処理によるホルモン分泌の変化 下垂体前葉には少なくとも5種類のホルモン産生細胞が含まれるが、その基礎分泌は2種類に大別される。1)基礎分泌能が高く、視床下部由来の抑制因子による調節を受ける群(GHとPRL)、2)基礎分泌能が低く、視床下部由来の促進因子による調節を受ける群(LH、FSH、TSH、ACTH)の2つである。第1章では、下垂体前葉初代培養細胞からのホルモン基礎分泌に対するトリプシン処理の影響を調べた。ラット下垂体前葉細胞をマイクロキャリア・ビーズ上で4日間培養し、その後、細胞表面潅流培養法に供し、潅流した培養液中の下垂体ホルモン濃度をRIAにより測定した。細胞を0.25%のトリプシンを含む培養液で5分間処理した結果、PRL分泌は未処理群に比べほぼ測定不能レベルまで低下したものの、120分後から分泌の回復がみられ、さらに100分後には完全に回復した。このPRL分泌の回復は蛋白質合成に依存している。他のホルモン分泌についても同様に調べた結果、GHにはPRL同様の基礎分泌低下が起こるものの、TSHとACTHは分泌が亢進し、LHとFSHはほとんど変化しなかったことから、PRLとGHの著しい分泌低下はトリプシン処理による非特異的な細胞傷害によるものではないことが分かった。次に、トリプシン処理した後、それぞれの下垂体ホルモンに対する分泌促進因子や高濃度のカリウムによる分泌刺激を与えたときのホルモン分泌を調べたところTSH、ACTH、LH、FSH分泌はそれぞれの分泌促進刺激に反応し分泌が亢進するのに対し、PRL・GH分泌は、全く分泌刺激に反応しなかった。これらの結果から、PRLとGHの基礎分泌には、トリプシン感受性細胞膜蛋白質(TSMPs)の発現が関与していることが示唆された。 第2章GH3細胞膜を抗原とした単クローン抗体作成の試み GH/PRL細胞系列特異的に発現するTSMPsを探索するために、GH3細胞精製膜画分を抗原として単クローン抗体の作成を試みた。ポリエチレングリコールによる細胞融合の結果、442ウェルのうち107個のウェルでハイブリドーマのコロニー形成が観察された。その全てにおいてGH3細胞を抗原としたELISAを行いGH3細胞に結合能を有するクローンを選択し、結合能の高い5つのクローンII-13、III-17、IV-16、IV-21、IV-24を選択した。その内の一つの単クローン抗体III-17(IgM.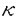 )の結合能は、GH3細胞をトリプシンで処理することにより低下した。III-17抗体を用いた免疫組織化学を行い正常下垂体組織を染色することにより、本抗原は全てのGH産生細胞と一部のPRL産生細胞に存在し、他のホルモン産生細胞には発現していないことが判明した。更に、本抗体を下垂体初代培養細胞に加え、GHとPRL分泌に対する影響を調べたところ、GH分泌は変化しないのに対して、PRL分泌を著しく亢進した。本抗体を用いたWestern Blottingの結果、還元状態でのみ66KDaの分子を認識したことから、本抗体の抗原決定基は蛋白質の高次構造の構築に重要なエピトープを認識していること、また、GH3細胞において存在が知られている既知の受容体に該当しない新規の分子であることが推察された。p66はGH/PRL細胞系列の分化及びPRL分泌に関わるTSMPsの候補分子と考えられる。 )の結合能は、GH3細胞をトリプシンで処理することにより低下した。III-17抗体を用いた免疫組織化学を行い正常下垂体組織を染色することにより、本抗原は全てのGH産生細胞と一部のPRL産生細胞に存在し、他のホルモン産生細胞には発現していないことが判明した。更に、本抗体を下垂体初代培養細胞に加え、GHとPRL分泌に対する影響を調べたところ、GH分泌は変化しないのに対して、PRL分泌を著しく亢進した。本抗体を用いたWestern Blottingの結果、還元状態でのみ66KDaの分子を認識したことから、本抗体の抗原決定基は蛋白質の高次構造の構築に重要なエピトープを認識していること、また、GH3細胞において存在が知られている既知の受容体に該当しない新規の分子であることが推察された。p66はGH/PRL細胞系列の分化及びPRL分泌に関わるTSMPsの候補分子と考えられる。 第3章GH3細胞の糖脂質組成 GH/PRL細胞系列の分化抗原として酸性糖脂質(ガングリオシド)の存在が示唆されている。しかし、下垂体前葉やGH3細胞のガングリオシド組成は解析されていない。そこで、GH3細胞より総脂質を抽出しガングリオシド画分を精製・構造解析を行った。粗精製したガングリオシドを薄層クロマトグラフィーにより成分を解析した結果、FG1とFG2の2種類のガングリオシドが主要成分であることがわかった。DEAE-Sephadex A-25カラムとTLC解析によって、FG1、FG2はN-アセチル-シアル酸を含むモノシアロガングリオシドであった。更に、高速原子衝撃法質量分析・核磁気共鳴を用いて構造解析を行ったところ、FG1とFG2はFucosyl GM1、 Gal-Fucosyl GM1と決定された。TLC免疫染色では、抗Fucosyl GM1抗体(CRD73-6)がFG1を、抗ヒトB型抗原決定基抗体がFG2を認識することが確認された。 Gal-Fucosyl GM1と決定された。TLC免疫染色では、抗Fucosyl GM1抗体(CRD73-6)がFG1を、抗ヒトB型抗原決定基抗体がFG2を認識することが確認された。 GH3細胞を1nM 17 -estradiol、10nM EGF、300nMインスリンを含む培地で6日間培養することにより、PRL分泌及び細胞内貯蔵量の増加・GH分泌の低下・接着性及び形態の変化を伴った分化を誘導することができる。GH3細胞を同条件で培養することによりFG1とFG2の発現は2倍以上増加した。FACS解析の結果、細胞表面上のFG1とFG2の発現は、分化前後で変化しないことがわかった。ホルモン存在下及び非存在下で培養した細胞を抗Fucosyl GM1抗体(CRD73-6)と抗ヒトB型抗原決定基抗体染色した結果、ホルモン非存在下では、細胞表面が優位に染色されるのに対し、ホルモン存在下では、細胞表面のみならず核の周囲ならびに核膜が強染された。また、正常下垂体細胞におけるFG1、FG2の発現をFACS解析した結果、それぞれ、27%,22%の細胞が陽性であった。これらの結果から、フコース含有ガングリオシドの細胞内局在性の変化がGH3細胞の分化に関係することが推察された。 -estradiol、10nM EGF、300nMインスリンを含む培地で6日間培養することにより、PRL分泌及び細胞内貯蔵量の増加・GH分泌の低下・接着性及び形態の変化を伴った分化を誘導することができる。GH3細胞を同条件で培養することによりFG1とFG2の発現は2倍以上増加した。FACS解析の結果、細胞表面上のFG1とFG2の発現は、分化前後で変化しないことがわかった。ホルモン存在下及び非存在下で培養した細胞を抗Fucosyl GM1抗体(CRD73-6)と抗ヒトB型抗原決定基抗体染色した結果、ホルモン非存在下では、細胞表面が優位に染色されるのに対し、ホルモン存在下では、細胞表面のみならず核の周囲ならびに核膜が強染された。また、正常下垂体細胞におけるFG1、FG2の発現をFACS解析した結果、それぞれ、27%,22%の細胞が陽性であった。これらの結果から、フコース含有ガングリオシドの細胞内局在性の変化がGH3細胞の分化に関係することが推察された。 以上、本研究ではGH/PRL細胞系列の分化抗原として3種類の分子の同定及び性状解析を行った。まずはじめに、下垂体前葉細胞をトリプシン処理すると、細胞種特異的にホルモン分泌が変化することを示した。特に、GHとPRLの顕著な分泌低下が起こった。次に、GH/PRL細胞系列特異的な細胞表面分子を探索し、GH細胞に発現しPRL分泌に関わる単クローン抗体III-17抗原(p66)とPRL細胞分化に伴い細胞内で発現が増加するフコース含有ガングリオシド(Fucosyl GM1,Gal Fucosyl GM1)を同定した。 これら細胞種特異的に発現する細胞表面分子群の解析を通して、下垂体前葉細胞の分化を新しい角度から捉えられるとともに、解放分泌機構における膜タンパク質の役割・シグナル分子としての糖脂質の機能等、より一般的な、細胞の分子機構の解明が期待できる。 |