現代医学の重要課題の一つは癌を解明し治療法を確立することである。癌細胞は高度に調節された細胞増殖制御の破綻により生じる。細胞の増殖制御は、細胞内外からの情報を受け、その情報に基づいて増殖の開始や停止を制御する細胞周期制御機構によって行なわれている。しかしながら、直接、哺乳類動物細胞の細胞周期制御機構を解明することは、強力な実験手段がないため、非常に困難である。これに対し、酵母は、はるかに分子遺伝学的取り扱いが容易な実験モデル生物であり、近年の成果により、細胞周期を含む細胞の基本制御システムが、酵母から哺乳類まで極めてよく保存されていることが明らかとなっている。そこで、私は、酵母の中でもより高等真核生物に似ている分裂酵母Schizosaccharomyces pombeを利用し、細胞周期制御の基本機構の解明を目指した。 細胞はDNA合成(S期)有糸分裂(M期)を繰り返し増殖する。その間を間隙(G1期、G2期)といい、ここでは極めて重要な細胞周期制御が行なわれる。G1期では、S期へ進行するか、分化するか、静止するかの決定が行なわれ、G2期ではM期へ進行するタイミングが決定されている。 分裂酵母は、細胞周期制御機構の普遍的な面を理解するための優れたモデル生物であが、最近の著しい進歩にかかわらず、その制御機構は依然として未知の部分が多い。細胞周期の開始に関わる新規制御因子を単離するために、私は、温度感受性cdc10-129またはcdc24-M38変異株の多コピー抑圧遺伝子をスクリーニングした。Cdc10はRes1/2及びRep1/2とともに、S期の開始に必要な転写因子複合体を構成する。cdc24+遺伝子は未同定であり、その変異株はS期の進行に欠損を持つ。スクリーニングの結果、cdc10-129変異株の多コピー抑圧遺伝子tbh1+、cdc24-M38変異株の多コピー抑圧遺伝子4種類とcdc24+遺伝子と考えられる遺伝子を単離することができた。 cdc10-129変異株の多コピー抑圧遺伝子tbh1+は、DNA複製開始点に働いて複製開始に必須なMCMファミリーの一つの変異株であるcdc19-P1変異株も多コピーで抑圧できた。tbh1+のcDNAの塩基配列を決定したところ、一つのORFが存在し、438アミノ酸の蛋白質Tbh1をコードしていた。Tbh1は分子量約49,000の蛋白質であり、N末にロイシンジッパー構造と中央部にATP/GTP結合部位モチーフを持っていた。データベースの検索により、Tbh1はヒトのtat-binding protein及び出芽酵母のYTA1遺伝子産物とアミノ酸配列で約70%という極めて高い相同性を持ち、その分裂酵母ホモログと考えられた。tat-binding proteinはhuman immunodeficiency virus(HIV)のtatと結合する蛋白質として発見され、転写因子として考えられてきたが、最近、26Sプロテアソームの構成要素でもある可能性が提唱されている蛋白質である。26Sプロテアソームはポリユビキチン化された蛋白質を特異的に分解する巨大なプロテアーゼ複合体である。 tbh1+をura4+に置き換えることにより、tbh1破壊株を作製した。tbh1破壊細胞は、発芽するが、一度も細胞分裂を完了せず致死であった。次に、フローサイトメトリーによる解析やDAPI染色による核や、抗チュブリン抗体によるスピンドルを観察した結果、tbh1破壊細胞は、S期の開始が約2時間遅れるもののDNA合成は完了し有糸分裂の中期に停止していることが明らかになった。 tat-binding proteinは26Sプロテアソームの構成要素である可能性が提唱されているため、その可能性を検討した。もし、Tbh1が26Sプロテアソームの構成要素ならばtbh1破壊細胞はポリユビキチン化された蛋白質を分解できないはずである。tbh1破壊細胞の蛋白質を、抗ポリユビキチン抗体を用いたウエスタンブロティングで解析したところ、予想通り、ポリユビキチン化された蛋白質が異常に蓄積していることが明らかになった。この結果から、Tbh1さらにはtat-binding proteinも26Sプロテアソームの必須の構成要素であることが強く示唆された。 以上のことから、tbh1破壊細胞はポリユビキチン化された蛋白質の著しい蓄積を伴いながら、有糸分裂の中期で停止する。中期/終期移行にCut2とCdc13のユビキチン依存的な蛋白質分解が必要なので、Tbh1の生物学的役割は、26Sプロテアソームのサブユニットとして中期/終期移行時にこれらの蛋白質を分解することだと考えられる。さらに、tbh1+の過剰発現によりcdc10とcdc19変異株を抑圧できることと、その破壊株でS期の開始が少し遅れることは、tbh1+がG1/S期移行にも機能を持つことを示唆する。 cdc24-M38変異株は、制限温度で増殖を停止し、典型的なcdc表現形を示す。cdc24+遺伝子の作用点は遷移点解析により、S期であることが判明している。私は、最初に、対数増殖期または窒素源枯渇下のcdc24-M38変異株を許容温度から制限温度へ移し、フローサイトメトリーにより、DNA含量を解析した。cdc24-M38変異株は、S期の遅延を示さず、G2DNA含量で細胞周期を停止した。これは意外なことではなく、他のS期のcdc変異株でもよくみられる現象である。S期のcdc変異株はチェックポイント変異株のrad1変異株と二重変異株を作製すると、制限温度でS期完了前に異常なM期に進入する。同様にcdc24-M38 rad1二重変異株は制限温度にすると、他のS期のcdc変異株と同様に異常な細胞分裂を開始した。 次に、cdc24+遺伝子の機能を明らかにするために、cdc24-M38変異株を多コピーで抑圧する遺伝子をクローニングし、5種類の抑圧活性をもつ遺伝子を単離した。その一つは、この変異株を36℃でも抑圧できることからcdc24+遺伝子自身と考えられた。塩基配列を決定したところ、一つのORFが存在し、501アミノ酸、分子量約58,000の蛋白質をコードしていた。その蛋白質は、既知の蛋白質とは相同性を持たず、顕著なモチーフも存在しなかった。残りの4つの多コピー抑圧遺伝子の塩基配列を決定しホモロジー検索をしたところ、cdc24+遺伝子の機能を考える上での重要な情報が得られた。一つは既知の遺伝子pcn1+でありPCNA(proliferating cell nuclear antigen)をコードする。もう一つは新規遺伝子rfc1+である。rfc1+は487アミノ酸の蛋白質をコードし、その蛋白質はヒト及び出芽酵母のRF-C(replication factor-C)の大サブユニットと約40%の相同性を持ち、その分裂酵母ホモログであると考えられた。DNA鎖の伸長は、PCNAとRF-Cを構成する5個のサブユニット及びDNAポリメラーゼ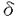 / /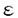 を含むDNA合成装置により行われる。それゆえ、pcn1+及びrfc1+が、cdc24-M38変異株を比較的強く抑圧することは、Cdc24がDNAポリメラーゼ/とPCNAとRF-Cを含むDNA合成装置の重要な構成要素であることを示唆している。 を含むDNA合成装置により行われる。それゆえ、pcn1+及びrfc1+が、cdc24-M38変異株を比較的強く抑圧することは、Cdc24がDNAポリメラーゼ/とPCNAとRF-Cを含むDNA合成装置の重要な構成要素であることを示唆している。 残りの一つは、ppa1+であることが塩基配列の決定により判明した。ppa1+は2A型脱リン酸化酵素をコードしている。このことは、S期の進行の制御がリン酸化によりなされている可能性を示唆している。 今後、Tbh1のG1/S期移行における標的蛋白質及びCdc24のDNA複製における機能を同定することによって、S期の開始及び進行の制御機構の解明に重要な手掛りが得られるものと期待される。 |