インターロイキン7(IL-7)はpre B細胞の増殖を支持する増殖因子として同定された糖タンパクである。その後、IL-7はpre B細胞のみならず未熟胸腺細胞に作用して増殖させることが判明した。さらにin vivoでのIL-7の機能を調べるために、抗IL-7ならびに抗IL-7レセプター(IL-7R)抗体をマウスに投与する実験が行われ、IL-7/IL-7Rシグナル伝達システムがB細胞ならびにT細胞の生成に重要であることが明らかになった。 そこで私は生体内におけるIL-7Rの機能を詳細に解明する目的で、IL-7R欠損マウスを作製しその解析を行った。その結果、IL-7RからのシグナルがT細胞およびB細胞の初期分化に重要であり、特に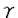 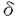 T細胞の分化もしくは維持に必須であることが明らかになった。IL-7がT細胞におよぼす作用として、(1)T細胞レセプター(TCR)鎖遺伝子のV-J組み換えを誘導すること、(2)T細胞に維持・増殖因子として働くことの2つが示唆されているが、IL-7R欠損マウスのT細胞におけるV-(D)-J組み換えについて解析したところIL-7RマウスではTCR鎖遺伝子のV-J組み換えが欠失していることが明らかになった。従って本研究により、IL-7RからのシグナルがTCR鎖遺伝子特異的なV-J組み換え誘導に必須であることが明らかになった。 T細胞の分化もしくは維持に必須であることが明らかになった。IL-7がT細胞におよぼす作用として、(1)T細胞レセプター(TCR)鎖遺伝子のV-J組み換えを誘導すること、(2)T細胞に維持・増殖因子として働くことの2つが示唆されているが、IL-7R欠損マウスのT細胞におけるV-(D)-J組み換えについて解析したところIL-7RマウスではTCR鎖遺伝子のV-J組み換えが欠失していることが明らかになった。従って本研究により、IL-7RからのシグナルがTCR鎖遺伝子特異的なV-J組み換え誘導に必須であることが明らかになった。 まず、ジーンターゲティング法によりIL-7R欠損マウスを作製した。129系マウス染色体ライブラリーより4IIL-7R染色体遺伝子を単離し、IL-7R第2エクソンをネオマイシン耐性遺伝子に置換するターゲティングベクターを構築した。このDNAをES細胞にエレクトロポレーション法によって導入し、145個の薬剤耐性クローンから2個の相同組み換え体をサザン法で同定した。これら2クローンのうち1クローンから作製したキメラマウスはES細胞の性質を子孫に伝達し、これらヘテロ接合体間の交配によりIL-7R欠損マウスを得た。IL-7R欠損マウスはメンデル比に従って得られ、同腹の野生型マウスとの間に外見上の差は認められなかった。IL-7Rの発現を調べたところ、IL-7R欠損マウスではエクソン2を欠損するmRNAを発現していたが、抗IL-7R単クローン抗体を用いてFACS解析したところ細胞表面へのIL-7Rの発現は認められなかった。 つぎに、IL-7R欠損マウスおけるリンパ球の分化をFACSを用いて解析した。IL-7R欠損マウスのリンパ球は正常マウスに比べ、骨髄では1/2に、胸腺では1/100に、脾臓では1/10程度に減少していた。B細胞ならびにT細胞の分化段階をFACSで解析したところ、骨髄のB細胞はHardyらの分類によるfraction AからBで、胸腺のT細胞はCD4-CD8-分画中のCD44+CD25-からCD44+CD25+の段階で分化が障害されていた。しかしながら、末梢にはB細胞ならびにT細胞が正常マウスの1/10程度ながらも存在しており、これらをLPSならびにCon Aで刺激したところ3H-チミジンの取り込みは正常であった。このことは、IL-7R欠損によってリンパ球の分化・増殖が重度に障害されるものの、末梢における増殖反応は阻害されないことを示唆する。さらにT細胞をFACSで解析したところ、胸腺、脾臓、腸管上皮、肝臓において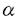 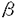 T細胞は存在するものの、T細胞は完全に欠失することが明らかになった。また皮膚上皮に分布するT細胞の一種であるDendritic Epidermal T Cells(DETC)も完全に欠失していた。一方、IL-7R欠損マウスではNK細胞は正常に存在し、NK活性についても野生型マウスとIL-7R欠損マウスとの間に著明な差は認められなかった。 T細胞は存在するものの、T細胞は完全に欠失することが明らかになった。また皮膚上皮に分布するT細胞の一種であるDendritic Epidermal T Cells(DETC)も完全に欠失していた。一方、IL-7R欠損マウスではNK細胞は正常に存在し、NK活性についても野生型マウスとIL-7R欠損マウスとの間に著明な差は認められなかった。 以上の結果から、IL-7Rからのシグナルはリンパ球の初期分化に重要であり特にT細胞の分化もしくは維持に必須であること、ならびにNK細胞の分化・増殖にはIL-7Rからのシグナルは必須でないことが明らかになった。 IL-7がT細胞に及ぼす作用として、TCR鎖遺伝子のV-J組み換えを誘導すること、ならびにT細胞の維持・増殖因子として働くことが2つ示唆されている。そこでこの問題を明らかにするために私はIL-7R欠損マウスのTCR遺伝子のV-(D)-J組み換えについて解析した。 まずIL-7Rヘテロ接合体ならびにIL-7R欠損マウスの胸腺細胞からDNAを抽出し、これらマウスの胸腺細胞DNAにおけるV-(D)-J組み換えをJ1,J2,J1,J1プローブを用いてサザン解析した。その結果、IL-7Rヘテロ接合体マウスの胸腺細胞DNAではJ1とJ2locusの70%以上のalleleで組み換えが認められ、V1.2-J2ならびにV2-J1組み換え体が検出された。しかしながら、IL-7R欠損マウスではTCR鎖遺伝子のV1.2-J2とV2-J1組み換え、ならびにそれに伴うgermline fragmentの減少は検出できなかった。一方、J2,J1,J1プローブを用いたサザン解析では、TCR,,鎖遺伝子のV-(D)-J組み換えについて両者の間に著明な差は認められなかった。 さらに詳細に調べるために、IL-7R欠損マウスの胸腺細胞DNAにおけるV-(D)-J組み換えをTCR鎖、鎖、鎖遺伝子のプライマーを用いてPCR法により解析した。その結果、IL-7R欠損マウスではすべてのTCR鎖遺伝子のV-J組み換えが著明に減少していた。しかしながら、TCR鎖遺伝子のV-D-J組み換えについてはIL-7Rヘテロ接合体マウスとIL-7R欠損マウスとの間に著明な差は認められなかった。最近、fetal thymic organ cultureにおいてIL-7がTCR鎖遺伝子のD-J組み換えを誘導することが報告されていたが、本実験ではTCR鎖遺伝子のD-J組み換えならびにV-DJ組み換えについてIL-7Rヘテロ接合体マウスとIL-7R欠損マウスとの間に著明な差は認められなかった。 またV-D-J組み換えに関与するRag-1,Rag-2,Ku-80,TdT遺伝子の発現についてRT-PCR法により解析したところ、IL-7Rヘテロ接合体マウスとIL-7R欠損マウスとの間に著明な差は認められなかった。 以上の結果、IL-7RからのシグナルがTCR鎖遺伝子特異的なV-J組み換えを誘導すること、ならびにすべてのTCR鎖遺伝子のV-J組み換えに必須であることが明らかになった。従ってIL-7R欠損マウスがT細胞が欠失する原因の少なくとも1つは、TCR鎖遺伝子のV-J組み換えが誘導されないことであることが示唆された。 IL-7Rはcommon鎖とヘテロダイマーを構成する。また、このcommon鎖はIL-7Rのみならず、他のサイトカインレセプター(IL-2R、IL-4R、IL-9R、IL-15R)に共有される。IL-2、IL-4ならびに両者を欠損するマウスではリンパ球の初期分化に異常は認められないものの、common鎖を欠損するマウスではリンパ球の初期分化と末梢での増殖が障害され特にT細胞とNK細胞が欠失する。本研究で得られた結果から、リンパ球の初期分化とT細胞の分化もしくは維持にはIL-7R/common鎖からのシグナルが必須であり、末梢でのリンパ球の増殖反応とNK細胞の分化・維持にはIL-15R等のcommon鎖を共有する他のサイトカインレセプターからのシグナルが重要であることが示唆される。 また本研究によりIL-7RからのシグナルがTCR鎖遺伝子に特異的なV-J組み換えの誘導に必須であることが明らかになった。最近、免疫グロブリンやTCR遺伝子のlineageならびにstage特異的なV-(D)-J組み換えを調節する機構としてエンハンサーによる制御が示唆されている。従って、IL-7RからのシグナルがTCR鎖遺伝子のエンハンサーに作用してTCR鎖遺伝子に特異的なV-J組み換えを制御していることが考えられる。IL-7はT細胞においてSTAT5を活性化する。さらにTCR鎖遺伝子のエンハンサーにSTAT5結合部位が存在する。従ってIL-7Rからのシグナル伝達においてSTAT5がTCR鎖特異的なV-J組み換え制御に関与していることが考えられる。 IL-7R欠損マウスはサイトカインおよびサイトカインレセプター欠損マウスの中でもリンパ球の初期分化の障害、特にT細胞の消失が明瞭に認められる数少ないマウスである。従ってこのマウスと変異IL-7RやTCRトランスジェニックマウスとを交配することによりIL-7Rの生体内における機能を詳細に解析することが期待される。また本研究によりIL-7RからのシグナルがTCR鎖遺伝子特異的なV-J組み換えに必須であることが明らかになったが、この成果はTCR遺伝子座のみならず他の抗原レセプター遺伝子の高次組み換え機構を解明する糸口になると思われる。 |