本研究は、広範な生物種において免疫応答や炎症、発生などに関与しているとされる転写調節因子群Rel/NF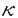 Bの制御機構を明らかにするため、Rel/NFBの制御蛋白質であるIB Bの制御機構を明らかにするため、Rel/NFBの制御蛋白質であるIB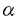 のリン酸化や分解を受ける機構の解析を試みたものである。第一章においてはin vitroでIBのリン酸化を行う活性の検出を、また第二章ではin vivoでIBのシグナルによる分解やリン酸化に必要とされるIB内の領域の決定をそれぞれ試みており、具体的に次のような結果を得ている。 のリン酸化や分解を受ける機構の解析を試みたものである。第一章においてはin vitroでIBのリン酸化を行う活性の検出を、また第二章ではin vivoでIBのシグナルによる分解やリン酸化に必要とされるIB内の領域の決定をそれぞれ試みており、具体的に次のような結果を得ている。 まず第一章に関しては 1.Jurkat細胞抽出液にGST-IB融合蛋白質を加えて抽出液中の蛋白質と十分混合した後にGST-IB融合蛋白質を沈殿させ、放射性ATPを用いたリン酸化反応を行ったところ、この融合蛋白質に対して強い放射活性の取り込みが見られ、抽出液中のキナーゼ活性によるリン酸化と考えられた。またこの活性はGST蛋白質による前処理で影響されないがGST-IB融合蛋白質による前処理で活性を吸収できることから融合蛋白質のIB部分に結合していることが分かった。更にリン酸化した融合蛋白質をトロンビン処理することにより融合蛋白質のIB部分がリン酸化されている事も確認された。従ってこの活性はIBに結合し、かつIBをリン酸化する活性であることが明らかとなった。 2.この活性は細胞に対するTNF処理の有無で変化が見られなかったことから細胞細胞外刺激に依存しておらず、またこのリン酸化によってGST-IBとRel蛋白質の結合力は影響を受けなかった。 3.このリン酸化は主にIB蛋白質の6番目のアンキリンリピートからカルボキシル末端の約70アミノ酸残基中のセリン・スレオニンに対して起こっていることが明らかとなった。 また、第二章ではIBミュータントを用いて実際に細胞内に於いてIB内のリン酸化或いは分解に必要となる部位を決定することを試みた結果から 4.HeLa細胞のIB恒常発現クローンを単離してTNF刺激を行うと、内在性のIBと同様の時間経過で外来性IBの分解が観察されたため、各種ミュータントIBの細胞外刺激に対する挙動を検討する実験系としてIBミュータントをHeLa細胞に導入した恒常発現クローンを用いることが有効であると判明した。 5.この実験系を用いて検討した結果、アミノ末端約70アミノ酸を欠くIBミュータントではシグナル依存的リン酸化・分解が共に起こらず、これら両方に必要な部位がこの領域に存在すると考えられた。また、IBの中央部に存在するアンキリンリピート領域はシグナル依存的リン酸化には必要ないもののシグナル依存的分解には必須であることが分かった。カルボキシル末端領域には蛋白質の分解に関与していると言われるPEST様配列が存在する事から、シグナル依存的分解への関与が期待されたが、このPEST様配列の殆ど全てを欠失したIBミュータントはTNF刺激によって内在性のIBと変わらないリン酸化・分解パターンを示し、IB蛋白質のPEST様配列がシグナルによるリン酸化や分解に関与していない事が明らかとなった。これらの結果から、IBのシグナル依存的リン酸化及び分解はそれぞれ異なる認識機構で制御されていることが判明した。 以上、本論文は転写調節因子群Rel/NFBの制御機構を、その制御蛋白質であるIBのリン酸化及び分解という観点から解析し、多くの情報を提供している。これらは未だ明らかとなっていないRel/NFB活性化に至る細胞内シグナル伝達機構の解明に貢献するものであり、学位の授与に値するものと考えられる。 |  Bの制御機構の解析
Bの制御機構の解析