様々な病原体の侵入を防ぐ免疫機構は、主にリンパ球によって制御されている。リンパ球は基本的に、T細胞とB細胞の2つのカテゴリーに分類することができる。B細胞は、抗体を分泌して病原体を不活化する、ないしは補体系と共役して破壊する液性免疫をつかさどる。T細胞は、いくつかのタイプに分かれて様々な機能を分担しており、一部のT細胞はB細胞の分化や抗体産生を調節し、マクロファージなどと共役して病原体を排除する機能を持つのでヘルパーT細胞と呼ばれる細胞集団が存在する。抗原提示細胞によって抗原を提示され最初に活性化されるリンパ球がヘルパーT細胞である。この細胞によってどの様な免疫担当細胞が活性化されるかによって、誘導される免疫応答も異なってくると考えられている。B細胞もその例外でなく、抗原受容体によって抗原を認識した細胞は、ヘルパーT細胞との相互作用によって活性化され、抗体産生細胞へと分化し、さらには抗体のクラスを換え最も効果的な抗体を分泌するようになる。しかしながら、抗原を認識したにも関わらず、ヘルパーT細胞との相互作用が無いと不活化されて、増殖を抑制されるか細胞死を起こして排除されると考えられている。ヘルパーT細胞との相互作用は、細胞間の直接の相互作用とサイトカインなどの因子を介した間接の相互作用があり、双方の情報の総和としてB細胞の活性化機構が機能する。直接相互作用で最も重要な情報をB細胞側に伝達すると考えられているのがCD40分子である。CD40に対するリガンド(CD40L)は活性化T細胞上に発現されることが明らかとなっている。血漿中にIgG、IgA、IgE、などのサブクラスの抗体がほとんど検出できなく、重度な免疫不全を引き起こす、高IgM血漿免疫不全症候群の患者はCD40Lの欠損あるいは変異によってCD40を介した情報伝達機構が不活化されていることが原因であることが明らかにされた。また、CD40のノックアウトマウスの解析から、二次リンパ組織における胚中心の不形成、クラススイッチの欠如など、ほとんどHIMの患者の症状を再現でき、これらのことから、CD40-CD40L機構は、T細胞依存性のB細胞の活性化機構に必須であると考えられるようになった。加えて、自己認識したB細胞の排除機構のモデル系とした、抗原受容体を介した情報によるB細胞細胞死の誘導系に、CD40を介した情報を加えると、その細胞死が抑制されたことから、B細胞の選択的増殖にもCD40-CD40L機構が深く関与していると考えられるようになってきた。私は、CD40を介した情報伝達系の解析が、B細胞の活性化機構と自己認識した細胞の排除機構の解明につながると考え、免疫学の大きな課題の一端を解明することができるのではないかと考えた。 CD40は、TNF受容体(TNFR)ファミリーに属する、分子量約50kDの細胞膜を一回貫通するI型の膜糖蛋白質である。このファミリーに属する受容体には、CD40の他、1型TNF受容体(TNFR-1)、2型TNF受容体(TNFR-2)、Fas抗原などが含まれる。特に、TNFR-1とFas抗原は細胞死を媒介する受容体として知られるが、その細胞内領域にはCD40の細胞内領域と有為な相同性を示す領域があり、TNFR-1、Fasの両受容体とCD40の情報伝達系に共通する機構があるのではないかと予想された。しかしながら、CD40の細胞内領域に結合するとしてクローニングされた分子はTNFR-2に結合するとしてクローニングされたTRAFファミリーに属する分子で、TNFR-1やFas抗原の細胞内領域に結合するとしてクローニングされたDeathドメインを持つ情報伝達因子群とは異なるものであった。 B細胞の活性化と分化においてその重要性は上述のように知られているCD40であるがその情報伝達機構、特に細胞死抑制の情報伝達機構の解析は私が初めて焦点を当てたと言える。私はまず、CD40によるB細胞の細胞死抑制の分子機構を明らかにした。 マウスB細胞株WEHI-231細胞にヒトCD40の恒常発現株を樹立し、ヒトCD40を抗体によって架橋することによりその情報伝達系を活性化できるようにした。図1に示すように、CD40を介した情報により、bcl-x遺伝子の発現が誘導され、細胞死を誘導するsIgMからの情報により、強く抑制されることが明らかとなった。 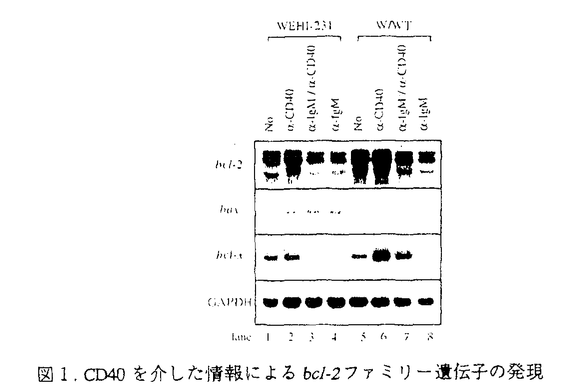 図1.CD40を介した情報によるbcl-2ファミリー遺伝子の発現 図1.CD40を介した情報によるbcl-2ファミリー遺伝子の発現 以上の結果より、CD40を介した細胞死抑制活性とbcl-x遺伝子の発現誘導とに相関性が認められた。bcl-x遺伝子にはスプライシングの異なる2つのmRNAが存在し、CD40によって発現が誘導されるのは細胞死の抑制活性を示すBcl-xLであったことから、細胞死抑制の効果はBcl-xLによってもたらされることが強く示唆された。次に、Bcl-xLの恒常発現細胞をWEHI-231細胞に樹立し、sIgM架橋によって細胞死が誘導されるかを検討した。この細胞の細胞死は抑制されたが、G1期で細胞周期が停止することは解除できずに増殖できないことが明らかとなった。そこで、sIgM架橋後のサイクリン依存性キナーゼ、Cdk4,6に注目し、その蛋白量を検討した。図2に示すように、sIgM架橋後24時間でCdk4,6共に蛋白量の減少が認められ、CD40を介した情報によってその減少は抑制されることが分かった。 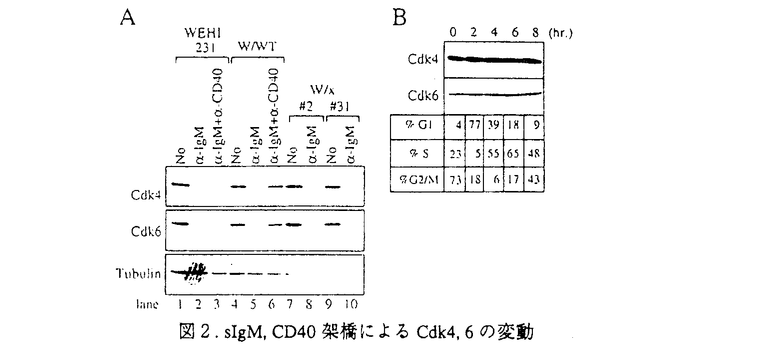 図2.sIgM,CD40架橋によるCdk4,6の変動 図2.sIgM,CD40架橋によるCdk4,6の変動 図2、Bでは細胞周期を同調させ、各周期でCdk4,6の蛋白量に変化がないことを確認している。 以上の結果から、CD40を介した細胞死抑制に関わる情報は、細胞死そのものの抑制(Bcl-xL)と細胞周期再開(Cdk4,6)という現象を引き起こし、その協調によって細胞死が抑制されるのだろうと考えられた。 次に、CD40を介した情報伝達因子のクローニングを試みた。Yeast Two Hybrid法を用いて、CD40細胞内領域に会合する分子をクローニングした。多くは、これまで報告されている、TRAF2,TRAF3であったが、新規のTRAFファミリーに属する、2つの遺伝子を得ることができた。それぞれ、5番目、6番目のTRAFファミリーであるので、TRAF5,TRAF6と命名した。表1にTRAF5,TRAF6のTRAFファミリー間に見られるTRAF-C domainにおける相同性を示した。 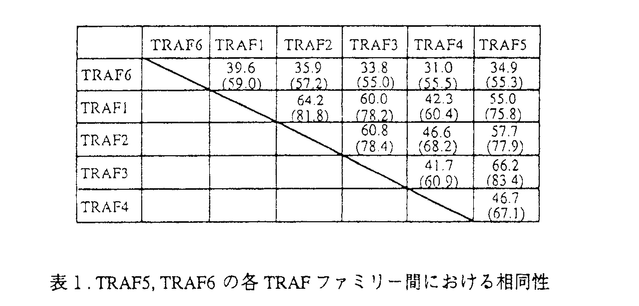 表1.TRAF5,TRAF6の各TRAFファミリー間における相同性 表1.TRAF5,TRAF6の各TRAFファミリー間における相同性 TRAF5は、アミノ酸配列上TRAF3に最も高い相同性を示し、CD40の細胞内領域246-269アミノ酸の領域に会合することが示された。この領域は、CD40の細胞死抑制の情報伝達に必要な領域に一致する。また、単独で細胞に過剰発現させるとNFkBの活性化を誘導し(図3)、N末の領域を欠損させた変異体は、CD40によるCD23の発現誘導を阻害したことから、CD40の情報伝達因子であると考えられた。 TRAF6は、アミノ酸配列上TRAF2と最も高い相同性を示したが、これまで報告された5種のTRAFファミリーのうちでその相同性は最も低い。CD40の細胞内領域の230-245アミノ酸領域に会合し、TRAF2,3,5とは異なる領域に会合することが示された。TRAF6もまた単独で過剰発現させることにより、NFkBの活性化を誘導し(図3)、またN末を欠損させたTRAF6はCD40△246(TRAF2,3,5の会合が認められないCD40の変異体で、NFkBの活性化が誘導される)のNFkBの活性化を阻害したことからCD40の情報伝達因子であると考えられた。 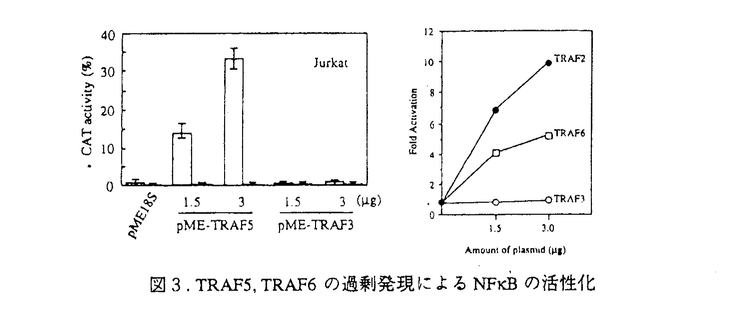 図3.TRAF5,TRAF6の過剰発現によるNFkBの活性化 図3.TRAF5,TRAF6の過剰発現によるNFkBの活性化 以上の結果から、クローニングされた新規TRAFファミリー、TRAF5,TRAF6はCD40の情報伝達因子であると考えられた。 今後、これらCD40に会合するTRAFファミリーがCD40が媒介する情報にどの様に関わっているのか解析していく必要があると考えている。 |