インターロイキン-5(Interleukin-5,IL-5)は、B細胞や好酸球に作用し増殖や分化を誘導するサイトカインである。また好塩基球にも作用し、ヒスタミン分泌を促すなど生体内の免疫応答や炎症反応を制御する。IL-5レセプター(IL-5R)はIL-5と特異的に結合する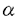 鎖と、単独ではIL-5と結合しないが鎖と共に機能的なIL-5Rを形成する 鎖と、単独ではIL-5と結合しないが鎖と共に機能的なIL-5Rを形成する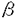 鎖より構築される。鎖はIL-5Rのみならず、IL-3レセプター(IL-3R)およびGM-CSFレセプター(GM-CSFR)の鎖としても機能し、これらのレセプターを介するシグナル伝達に必須である。IL-5の生理活性を理解するためには、IL-5Rを介するシグナル伝達機構を明らかにすることが重要である。 鎖より構築される。鎖はIL-5Rのみならず、IL-3レセプター(IL-3R)およびGM-CSFレセプター(GM-CSFR)の鎖としても機能し、これらのレセプターを介するシグナル伝達に必須である。IL-5の生理活性を理解するためには、IL-5Rを介するシグナル伝達機構を明らかにすることが重要である。 本研究では、IL-5によって誘導される増殖シグナルの伝達機構を解明することを目的とし、(1)マウスIL-5R鎖の細胞内領域のシグナル伝達における役割の解明とその機能領域を同定、(2)IL-5R鎖に対する抗体を用いてIL-5R鎖と会合する分子の検索、および(3)IL-5依存性を失ったIL-5非依存性増殖株の増殖機構の解析を行い、以下のような知見を得た。 (1)IL-5R鎖細胞内領域の機能および機能領域の同定 IL-5に特異的な結合を示すマウスIL-5R鎖(以下鎖と略す)は比較的短い細胞内領域(55アミノ酸残基)しか持たないが、細胞内領域を全て欠失させた場合IL-5シグナルの伝達機能を失うことが報告され、鎖もシグナル伝達に関与することが示された。そこで、鎖の細胞内領域のシグナル伝達における機能およびその機能領域を同定するため、種々の欠失変異をもつ鎖を作製し検討した。まず、PCR法を用いて細胞内領域の一部(6アミノ酸残基)を欠失した4種の変異鎖cDNAを作製しマウス鎖を内在的に発現するIL-3依存性骨髄前駆細胞株FDC-P1に遺伝子導入した。いずれの変異鎖を発現した細胞も高親和性IL-5Rを発現し、それらの解離定数は正常な鎖を発現した細胞の解離定数とほぼ同じであった。C末端側に欠失を持つ鎖を発現した細胞はIL-5に増殖応答を示したが、膜貫通領域直下のプロリンに富む領域を欠失した鎖を発現した細胞はIL-5に増殖応答を示さなかった。また、プロリンに富む領域を欠失した鎖を発現した細胞では、c-fos、c-jun、c-mycの発現および細胞内蛋白のチロシンリン酸化の誘導も同様に見られなかった。そこで、シグナル伝達に重要だと考えられるこのプロリンに富む領域のどのプロリン残基が大切であるかを調べるために点変異を持つ鎖を作製しFDC-P1に遺伝子導入した。プロリンに富む領域の3つのプロリンを全てアラニンに置換した鎖を発現した細胞ではIL-5に増殖応答を示さなかった。両端のプロリンをアラニンに置換した鎖を発現した細胞ではIL-5に弱く増殖応答を示した。 チロシンキナーゼ型レセプターやサイトカインレセプターで、レセプターのダイマリゼーションが活性化に重要であることが報告されている。IL-5Rの場合もレセプターの活性化とシグナル伝達のために鎖細胞内ドメインのダイマリゼーションのみで十分なのかを検討するため、細胞外と膜貫通部分が鎖、細胞内が鎖からなるキメラレセプターを作製して調べた。このキメラレセプターを発現している細胞はIL-5に応答して増殖反応を示した。このことにより、増殖シグナルは鎖細胞内ドメインのダイマリゼーションにより十分伝達されうることが示唆された。 以上よりIL-5の増殖応答には鎖細胞内領域のプロリンに富む領域が必要であるが、鎖で代行できうるため鎖は鎖のダイマリゼーションを補助している可能性が考えられた。 (2)サイトカインレセプターのシグナル伝達でJAKキナーゼと、その基質である転写因子STATの活性化が重要であることがいわれている。最近の研究によりIL-5のキメラレセプターを発現している細胞ではIL-5に応答してJAK1キナーゼ、STAT5の活性化は認められたものの野生株でみられるJAK2キナーゼの活性化は認められないことが共同研究者により報告された。そこでシグナル伝達に重要な鎖に会合する分子を同定し、それが鎖細胞内領域のどこに会合しているかを調べる目的で、鎖に対する抗体の作製を試みた。組み換えDNAの手法を用いて分泌型鎖を調製し、それをラットに免疫した。そのラットの脾臓細胞を用いてハイブリドーマを作製し、結合阻害実験によりIL-5と鎖の結合を阻害しない抗体を産生するハイブリドーマをスクリーニングした。得られた3種類の抗体を用いて免疫沈降を行ったところ、IL-5Rを発現している細胞のみから分子量60KDaのバンドが検出された。これによりこれらの抗体は鎖を認識し免疫沈降を行えるものとわかった。精製したこれらの抗体がIL-5によって誘導される細胞の増殖を阻害しないかを調べてみたところ、H7(IL-5中和活性を示す鎖に対する抗体)では2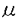 gの添加により増殖が完全に抑えられたのに対して、これらの抗体はその10倍量加えても約50%の増殖抑制を示すのみであった。また、多量のIL-5存在下でこれらの抗体が鎖を認識できるかを調べたところ、20nMIL-5の存在、非存在下では免疫沈降できる鎖量には変化が認められなかった。そのため、これらの抗体はIL-5のシグナルを伝えうる状態で鎖を免疫沈降できる可能性があると考えた。 gの添加により増殖が完全に抑えられたのに対して、これらの抗体はその10倍量加えても約50%の増殖抑制を示すのみであった。また、多量のIL-5存在下でこれらの抗体が鎖を認識できるかを調べたところ、20nMIL-5の存在、非存在下では免疫沈降できる鎖量には変化が認められなかった。そのため、これらの抗体はIL-5のシグナルを伝えうる状態で鎖を免疫沈降できる可能性があると考えた。 サイトカインレセプターを介するシグナル伝達にはチロシン残基のリン酸化が重要な役割を担っていることが知られており、IL-5の場合も鎖を含む数種類の細胞内タンパクがチロシンリン酸化される。そこで、今回作製した抗体を用いて免疫沈降実験を行い、リン酸化チロシンを含むタンパクが共沈されてくるかを調べた。IL-5依存性細胞株よりこれらの抗体で免疫沈降を行い、SDS-PAGEで展開後膜に転写し抗リン酸化チロシン抗体(4G10)でイムノブロットを行った。IL-5の刺激の有無にかかわらず、チロシンリン酸化された細胞内タンパクは認められなかった。鎖のリン酸化が認められなかったことからこれらの抗体がIL-5の中和活性を弱いながらも保持していることや、免疫沈降の条件により、鎖が解離した可能性もあることが類推された。これらの抗体を用いてシグナル伝達に重要な鎖に会合するリン酸化チロシンを含む分子を同定することはできなかった。 (3)IL-5依存性増殖株からのIL-5非依存性増殖株への変換機構の解析を行った。IL-5のシグナル伝達の解析には、IL-5応答性を持っていた細胞がIL-5応答性を失ったり、IL-5がなくても増殖するような変異株化細胞が得られれば有用である。そこで鎖を遺伝子導入しIL-5応答性を獲得した株化細胞をクローニングした後、IL-5非依存的に増殖するサブクローンを選択し、その増殖機構を調べた。IL-3依存性株化細胞で鎖の膜領域もしくは細胞外領域に変異が起こるとIL-3非依存的に増殖することが報告されているので、IL-5非依存性増殖株の鎖にも変異が起こった可能性を考えた。PCRにより膜近位の細胞外領域と膜貫通領域を増幅し調べたところ鎖に変異は認められなかった。このIL-5非依存性増殖株に抗鎖抗体や抗IL-5抗体を加えると、細胞増殖が抑制された。これよりこの細胞が培養液中に恒常的にIL-5を分泌している可能性が考えられた。そこでこの細胞の培養液中のIL-5の量を定量したところIL-5が検出された。また、IL-5mRNAの発現をノーザン法で調べると強いIL-5mRNAの発現がみられた。これらのことから、樹立したIL-5非依存性増殖株は自らIL-5を産生し、オートクリンループにより自立増殖をしていることがわかった。このIL-5非依存性増殖株が恒常的にIL-5を産生する機構を調べる一つの方法として、IL-5染色体遺伝子の構造をサザンブロット法により解析した。その結果、IL-5遺伝子上流側に正常マウス染色体には認められない新しい制限酵素切断部位が検出され、IL-5染色体遺伝子に構造的な変化が生じていることが推定された。 本研究により、IL-5のシグナルを下流に伝えるには、シグナル伝達に重要な役割を果たす鎖のみならず鎖の細胞内領域も必須であることが初めてわかった。細胞内ドメインの欠失変異の実験により鎖の細胞膜直下のプロリンに富む領域が増殖シグナルに重要であることも明らかにできた。しかし、IL-5に特異的なシグナル伝達系が本当に存在するのか、もし存在するのならばそれは鎖の細胞内のどの領域を必要としているのかはまだ未解明である。また、本研究では増殖シグナルを指標として調べてきたが、IL-5は好酸球やB細胞の分化を誘導する重要なサイトカインであり、分化の誘導には増殖シグナル伝達系とは異なる経路を利用していることも考えられる。そのため、IL-5にのみ見出されるシグナル伝達系の探索することも大切であると考えられる。今回IL-5刺激を完全に中和しない鎖に対する抗体を作製することができなかったが、IL-5に特異的なシグナルを調べるには、鎖を認識する抗体もしくはそれに代わる系の確立が必須であると考えられる。また、IL-5非依存性増殖株においては、変異がIL-5染色体遺伝子のみなのか、細胞内のIL-5シグナル伝達経路に異常が起こってないか検討の余地が残っていると思われる。 | 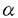 鎖の構造と機構の解析
鎖の構造と機構の解析