| (研究の目的) インターロイキン5(IL-5)はB細胞への抗体産生の誘導のほか好酸球の産生誘導、維持などの複数の作用を有している。IL-5に対する受容体は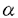 鎖と 鎖と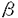 鎖から構成されている。IL-5受容体の特徴として鎖がIL-5に特異的であるのに対し、鎖はIL-3受容体および顆粒球マクロファージコロニー刺激因子(GM-CSF)受容体の鎖としても共有されていることが挙げられる。IL-5、IL-3およびGM-CSFは一部の細胞に対して共通の働きをもつが、これは共通の鎖(c)により同じ細胞内シグナル伝達経路が活性化されるためであると推測され、実際にcの細胞内ドメインの特定の領域がシグナル伝達に不可欠であることが報告されている。一方IL-5受容体鎖の細胞内領域もまたシグナル伝達に必要であることが示されているが、実際にどのような機能をもっているかについては知られておらず、IL-3受容体鎖およびGM-CSF受容体鎖との違いの有無を含めて注目される。IL-5特有の作用のひとつとしてB細胞初期発生への関与を示唆するいくつかの報告がなされている。つまり、マウスの骨髄細胞からIL-5依存性のCD5陽性プレB細胞株が樹立できること、IL-5遺伝子導入マウスではCD5陽牲B細胞亜集団が増加すること、IL-5受容体遺伝子欠損マウスではCD5陽性B細胞亜集団が減少することからIL-5はCD5陽性B細胞亜集団の発生に関与すると考えられる。しかし、それにどのようなシグナル伝達機構が関与しているかについては不明である。 鎖から構成されている。IL-5受容体の特徴として鎖がIL-5に特異的であるのに対し、鎖はIL-3受容体および顆粒球マクロファージコロニー刺激因子(GM-CSF)受容体の鎖としても共有されていることが挙げられる。IL-5、IL-3およびGM-CSFは一部の細胞に対して共通の働きをもつが、これは共通の鎖(c)により同じ細胞内シグナル伝達経路が活性化されるためであると推測され、実際にcの細胞内ドメインの特定の領域がシグナル伝達に不可欠であることが報告されている。一方IL-5受容体鎖の細胞内領域もまたシグナル伝達に必要であることが示されているが、実際にどのような機能をもっているかについては知られておらず、IL-3受容体鎖およびGM-CSF受容体鎖との違いの有無を含めて注目される。IL-5特有の作用のひとつとしてB細胞初期発生への関与を示唆するいくつかの報告がなされている。つまり、マウスの骨髄細胞からIL-5依存性のCD5陽性プレB細胞株が樹立できること、IL-5遺伝子導入マウスではCD5陽牲B細胞亜集団が増加すること、IL-5受容体遺伝子欠損マウスではCD5陽性B細胞亜集団が減少することからIL-5はCD5陽性B細胞亜集団の発生に関与すると考えられる。しかし、それにどのようなシグナル伝達機構が関与しているかについては不明である。 本研究ではB細胞初期分化におけるIL-5のシグナル伝達機構を解明することを目的として以下のような実験を行った。 1.IL-5刺激時にチロシンリン酸化を受ける細胞内分子について受容体鎖を共有するサイトカインであるIL-3およびGM-CSF、異なる受容体を用いるIL-2と比較し、IL-5特異的な、あるいは他のサイトカインと共通の経路の同定を試みた。 2.IL-5受容体鎖細胞内領域の細胞増殖に必須の領域として細胞膜直下のプロリンに富む領域が知られている。IL-5受容体鎖のシグナル伝達における機能を解明するためにこの領域に変異を加えた受容体を用いてシグナル伝達に与える影響を検討した。また、IL-5受容体がIL-6受容体などと同様に受容体鎖の重合によりシグナルを伝え得るかどうか検討するために細胞外が鎖、細胞内が鎖からなるキメラ受容体を用い、活性化されるシグナル伝達分子を野生型IL-5受容体の場合と比較した。 3.X染色体連鎖免疫不全(XID)マウスはCD5陽性B細胞亜集団の欠損を特徴とするB細胞分化増殖異常がみられ、一部の胸腺非依存性抗原に対する応答不全と血中IgM,IgG3の低値を示す。XIDマウスのB細胞はIL-5刺激による抗体産生の誘導に応答不全を示すことからそのB細胞分化増殖異常にもIL-5受容体を介するシグナル伝達の異常が関与していることが考えられる。骨髄長期培養の系を用いてB細胞初期分化におけるXIDマウスのIL-5応答性を検討し、さらにXIDマウス由来早期B細胞株でIL-5刺激によりチロシンリン酸化されるシグナル伝達分子を野生型の場合と比較した。 (方法) 1.IL-5応答性早期B細胞株Y16(IL-3にも応答する)にGM-CSF受容体鎖を遺伝子導入により発現させY16-GMRを得た。Y16-GMRをIL-3、IL-5およびGM-CSFで刺激しその細胞可溶化物からJAK1、JAK2、JAK3、SHC、Vav、STAT5の各分子を免疫沈降法にて精製しウエスタンブロットにてそのチロシンリン酸化を検出した。-カゼインプロモーター領域のプローブを用いたゲルシフト法にてSTAT5の活性化を検討した。IL-2応答性T細胞株CTLL-2にIL-5受容体鎖、鎖を遺伝子導入により発現させたCTLL5Rを用い、IL-2あるいはIL-5で刺激した際のJAK1,JAK2,JAK3のチロシンリン酸化を同様に検討した。 2.変異IL-5受容体鎖の機能解析 IL-5受容体鎖の第352アミノ酸残基から第355残基にみられるプロリン-プロリン-バリン-プロリン(ppvpと表記)部分の2つあるいは3つのプロリンをアラニン(A)に置換した変異受容体(pAvA,AAvp,pAvA,AAvA)および第357残基のプロリンをアラニンに置換した変異受容体(ppvpaA,ppvAaA)、さらに細胞外がIL-5受容体鎖、細胞内が鎖からなるキメラ受容体をIL-3依存性骨髄球系前駆細胞FDC-P1に遺伝子導入により発現させた。各受容体を発現した細胞についてJAK1,JAK2のチロシンリン酸化、STAT5の活性化を1.に準じて解析した。 3.XIDマウス由来IL-5依存性プレB細胞株の解析 3週齢のC57BL/6マウスおよび同xidマウスの骨髄細胞をWhitlock-Witte法にて培養し、さらに支持細胞株ST2およびIL-5存在下に限界希釈法にて株化した。細胞株表面分子は蛍光抗体染色後フローサイトメトリーにて解析した。IL-5刺激後のJAK1,JAK2,JAK3,STAT5およびSHCのチロシンリン酸化は1.に準じて解析した。またIL-5刺激後の細胞可溶化物からBtkを免疫沈降し、試験管内リン酸化反応によりその活性を測定した。 (結果および考察) 1.Y16-GMRにおいてはIL-3、IL-5、GM-CSFの各刺激後にJAK1、JAK2、STAT5、SHCおよびVavのチロシンリン酸化が共通に観察された。またゲルシフト法により各刺激後にSTAT5の活性化が同様に観察された。これよりJAK1,2-STAT5経路とSHCおよびVavを経由するRas-MAPK経路がIL-3,IL-5,GM-CSFに共通の細胞内シグナル伝達経路として働きうる可能性が示された。CTLL-5RにおいてはIL-2刺激時にはJAK1,JAK3が、IL-5刺激時にはJAK1,JAK2がそれぞれチロシンリン酸化され、IL-2およびIL-5刺激により異なる経路を介して細胞増殖が起こり得る可能性が示された。 2.IL-5刺激によりpAvAおよびAAvp変異体は野生型と同様にJAK1、JAK2のチロシンリン酸化およびSTAT5の活性化を誘導できたが、ApvA変異体ではこれらの変化は減弱し、AAvA変異体では全くみられなかった。これによりPPXP域が既存のプロリンに富むモチーフであるボックス1(PXP)およびSH3結合モチーフ(PXXP)とは異なる機能をもつことが示唆された。さらに、ppvAaA変異体においてもJAK1、2のチロシンリン酸化、STAT5の活性化は弱いながら認められたことから第355,357残基のプロリンがPXPモチーフをとして働く可能性は否定された。また、キメラ受容体を発現した細胞ではJAK1のチロシンリン酸化およびSTAT5の活性化は誘導されたが、JAK2のチロシンリン酸化はみられなかった。これより鎖の重合だけでは完全なシグナル伝達が起こらず、JAK2のチロシンリン酸化にはIL-5受容体鎖細胞内領域が重要な働きをもっていることが明らかになった。 3.XIDマウスからIL-5依存性の早期B細胞株が確立され、その頻度は野生型マウスを用いたときと変わらなかった。これはXIDマウスにおいても早期B細胞はIL-5に対して応答できる可能性を示すものである。XIDマウス由来の株は野生型由来のものと比べCD5抗原陽性の株の割合が低い傾向がみられた。このことからXIDマウスにおけるCD5陽性B細胞集団の欠損にIL-5応答不全が一部関与することが示唆された。XID由来IL-5依存性プレB細胞株(VpreB,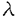 5陽性)をIL-5刺激するとJAK1、JAK2、STAT5、SHCのチロシンリン酸化が誘導された。また、XID由来、野生型由来の株ともIL-5刺激後のBtkの活性には変化がみらなかった。これらの結果よりXIDマウスにおいて少なくともプレB細胞まではIL-5による増殖シグナルは障害されていないことが明らかになった。また、プレB細胞におけるIL-5による増殖シグナルにはJAK1,JAK2-STAT5経路、あるいはSHCを介するシグナル伝達経路が使われ、XIDで変異のみられるBtkはシグナル伝達に関与していない可能性が示唆された。結論としてIL-5受容体を介するシグナルにはBtkを介する経路と介さない経路があり、プレB細胞では後者が使われていることが推測された。そのような経路の候補としてJAK1,JAK2-STAT5系路およびSHCを介するシグナル伝達経路が考えられた。 5陽性)をIL-5刺激するとJAK1、JAK2、STAT5、SHCのチロシンリン酸化が誘導された。また、XID由来、野生型由来の株ともIL-5刺激後のBtkの活性には変化がみらなかった。これらの結果よりXIDマウスにおいて少なくともプレB細胞まではIL-5による増殖シグナルは障害されていないことが明らかになった。また、プレB細胞におけるIL-5による増殖シグナルにはJAK1,JAK2-STAT5経路、あるいはSHCを介するシグナル伝達経路が使われ、XIDで変異のみられるBtkはシグナル伝達に関与していない可能性が示唆された。結論としてIL-5受容体を介するシグナルにはBtkを介する経路と介さない経路があり、プレB細胞では後者が使われていることが推測された。そのような経路の候補としてJAK1,JAK2-STAT5系路およびSHCを介するシグナル伝達経路が考えられた。 (今後の展望) この研究の結果IL-5のシグナル伝達経路において複数の経路が存在することが示唆され、それらが細胞の分化段階によって使い分けられている可能性が示された。今後それぞれの経路の構成分子を解明し、また、その使い分けの起こる機構を明らかにすることでB細胞分化増殖の制御機構の一端が明らかになると思われる。 |