本研究は、ラット頚動脈内膜剥離モデルを用い、in vivoの増殖期血管平滑筋で、平滑筋収縮において重要な役割を演じていると考えられるイノシトール1,4,5・三リン酸受容体(IP3R)の発現を免疫組織化学染色によって解析したものであり、下記の結果を得ている。 1.内膜剥離後早期(7、14日後)には、新生内膜の血管平滑筋で、IP3Rの発現が強く、特に管腔に面した平滑筋で最も強い発現が見られた。一方、この時点の中膜平滑筋では発現が低下していた。28日後には新生内膜管腔側でやや強い発現が見られたが、56日後には新生内膜、中膜全層に亙って均質で、正常対照血管の中膜とほぼ等しかった。 2.Proliferating cell nuclear antigen(PCNA)陽性の増殖細胞は内膜剥離7、14日後の新生内膜に多く見られ、特に管腔に面した細胞の殆どがPCNA陽性であった。即ち、PCNA陽性の増殖期血管平滑筋細胞とIP3Rを強く発現する細胞の分布が一致していた。以上より、IP3Rの発現が平滑筋増殖と密接に関係することが示された。 3.新生内膜管腔側でIP3Rの発現が高度であったのと対照的に、内膜剥離後早期の中膜では発現がむしろ低下していたが、これらの細胞は電子顕微鏡による形態観察ではいずれも合成型平滑筋であった。合成型平滑筋には更に細かい表現型の違いがあることがIP3Rの発現からも示された。 3.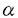 -平滑筋アクチンは全時期を通して、新生内膜、中膜ともにほぼ均質な発現を認め、IP3Rの発現と著しい違いを認めた。 -平滑筋アクチンは全時期を通して、新生内膜、中膜ともにほぼ均質な発現を認め、IP3Rの発現と著しい違いを認めた。 4.免疫電子顕微鏡では、内膜剥離14日後の新生内膜管腔側に、細胞質が網目状に濃染される血管平滑筋が観察された。これは透過型電子顕微鏡で観察された細胞内に網目状に発達した粗面小胞体の分布に一致していた。増殖期血管平滑筋におけるIP3Rの細胞内分布は、扁平な滑面小胞体に多いと報告されている小脳プルキンエ細胞における分布とは対照的に、粗面小胞体に多いことが示された。 以上、本論文はin vivoのラット頚動脈血管平滑筋において、平滑筋収縮において重要な役割を演じていると考えられるIP3Rの発現が、平滑筋増殖に依存して変化することを明らかにした。本研究は、動脈硬化病変部や血管形成術後の平滑筋増殖や平滑筋収縮異常の解明に重要な貢献をなすと考えられ、学位の授与に値するものと考えられる。 | 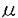 m厚の凍結切片とし、ヘマトキシリンーエオジン染色を行った。また、planimetryで新生内膜と中膜の面積及び内膜/中膜の面積比を求めた。(5)精製抗IP3R抗体、抗proliferationg cell nuclear antigen(PCNA)抗体、抗
m厚の凍結切片とし、ヘマトキシリンーエオジン染色を行った。また、planimetryで新生内膜と中膜の面積及び内膜/中膜の面積比を求めた。(5)精製抗IP3R抗体、抗proliferationg cell nuclear antigen(PCNA)抗体、抗