| 要旨 三量体GTP結合蛋白であるG蛋白の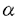 サブユニットである サブユニットである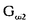 は組織特異的なプロトオンコジンであり、その点変異体のgip2には特定の細胞株に対して形質転換能があるが、その機序は不明である。今回の研究では早期反応遺伝子の一つであるc-fosのプロモーターがgip2による情報伝達の下流にあることを発見した。gip2はc-fosプロモーター上のSRE(serum responsive element)を介して正に制御し、RCE(retinoblastoma control element)を刺激して負に制御していた。このgip2のRCEに対する作用はレチノブラストーマ遺伝子蛋白(pRb)を介しており、gip2はpRbの発現の促進し、pRbを低リン酸化状態(活性化型)に制御する事の両方でpRbを活性化していた。gip2により形質転換することが知られているRat-1細胞株では、このgip2によるpRbを介した負の転写制御が欠失していた。かようにgip2はc-fosプロモーター上の2種類の領域を介してc-fosの転写を正負両方に制御しており、gip2による形質転換においてはpRbの活性制御が重要であると結論した。 は組織特異的なプロトオンコジンであり、その点変異体のgip2には特定の細胞株に対して形質転換能があるが、その機序は不明である。今回の研究では早期反応遺伝子の一つであるc-fosのプロモーターがgip2による情報伝達の下流にあることを発見した。gip2はc-fosプロモーター上のSRE(serum responsive element)を介して正に制御し、RCE(retinoblastoma control element)を刺激して負に制御していた。このgip2のRCEに対する作用はレチノブラストーマ遺伝子蛋白(pRb)を介しており、gip2はpRbの発現の促進し、pRbを低リン酸化状態(活性化型)に制御する事の両方でpRbを活性化していた。gip2により形質転換することが知られているRat-1細胞株では、このgip2によるpRbを介した負の転写制御が欠失していた。かようにgip2はc-fosプロモーター上の2種類の領域を介してc-fosの転写を正負両方に制御しており、gip2による形質転換においてはpRbの活性制御が重要であると結論した。 緒言 三量体GTP結合蛋白(G蛋白)は,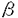 , ,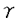 サブユニットで構成されており、受容体刺激の情報変換器として確立している。G蛋白の活性はグアニンヌクレオチドの結合により制御されている。G蛋白には内在性のGTPase活性があり、この活性でGTPをGDPに変換することにより自己を不活化させる。このG蛋白の中で、は増殖や形質転換を誘導する代表的なものである。の点変異であるQ205L、R179CとR179Hは内分泌腫瘍で発見されたが、これらは内在性のGTPase活性が欠失している変異であった。この活性が欠失した点変異体ではは活性化型の状態を保ち、依存性の情報伝達を活性化することで腫瘍形成している可能性がある。これらのgip2と呼ばれるの点変異体はRat-1線維芽細胞株に安定発現すると形質転換がみられる。かようにgip2は癌遺伝子であり、は前癌遺伝子である。 サブユニットで構成されており、受容体刺激の情報変換器として確立している。G蛋白の活性はグアニンヌクレオチドの結合により制御されている。G蛋白には内在性のGTPase活性があり、この活性でGTPをGDPに変換することにより自己を不活化させる。このG蛋白の中で、は増殖や形質転換を誘導する代表的なものである。の点変異であるQ205L、R179CとR179Hは内分泌腫瘍で発見されたが、これらは内在性のGTPase活性が欠失している変異であった。この活性が欠失した点変異体ではは活性化型の状態を保ち、依存性の情報伝達を活性化することで腫瘍形成している可能性がある。これらのgip2と呼ばれるの点変異体はRat-1線維芽細胞株に安定発現すると形質転換がみられる。かようにgip2は癌遺伝子であり、は前癌遺伝子である。 今回は2つの問題を提起した。第一にによる細胞増殖の機序の解明、第二にはgip2による形質転換能の組織特異性の解明である。これらの2つの問題は相互に関連している。今回対象としたc-fos遺伝子はc-jun等の早期反応遺伝子群の一つであり、前癌遺伝子(プロトオンコジン)である。AKR細胞ではTGF-b(Tranforming Growth Factor-)は早期反応遺伝子発現を百日咳毒素感受性G蛋白を介してに刺激する。又、G.L.Johnsonらのグループではgip2をRat-1細胞株に安定発現させると、MAPキナーゼが活性化され、c-junが発現する事を報告している。これらの研究から今回gip2によるc-fosプロモーター転写活性を解析した。 実験方法 cDNA発現プラスミド作成及びCATレポータープラスミド作成:活性化型G蛋白はgip1,gip2,gip3,gzp,gqp,gop,Ha-ras(V12G)を用いた。CATプラスミドはc-fosプロモーターCAT,TRE-CAT,CRE-CAT,c-mycプロモーターCAT,RCE-CAT,SRE-CAT,RbプロモーターCATを用いた。SRE部位を欠失したc-fosプロモーターCATプラスミド(c-fosdSRE-CAT)は4つのプライマーを用いたPCRで作成した。RCE部位を欠失したc-fosプロモーターCATプラスミドも同様に作成した。 細胞培養、cDNA形質導入と試料調整:細胞株はCHO細胞、Y79細胞、Rat-1細胞を用いた。径100-mm培養皿に1×106個の培養細胞をスプリットし、24時間後にgip2cDNAとCATcDNAをトランスフェクタムを用いて形質導入し、48時間後にCAT活性を測定した。 CATアッセイ:シンチレーション管内で[14C]ブチリルコエンザイムAと試料中のCATが反応した生成物である[14C]ブチリルクロラムフェニコールを反応液に重層したエコノフロアー-2に展開させ、その放射活性を反応開始200分後に測定した。CATの発現効率は共導入したヒトアルカリフォスファターゼ又はb-ガラクトシダーゼ活性で補正した。 G蛋白のADPリボシル化:試料を活性化した百日咳毒素により[32P]b-NAD存在下でADPリボシル化し、SDS-PAGE後、に相当する40-kDaのシグナルを測定した。 ウエスタンブロッテイング:試料を10%ポリアクリルアミドゲルSDS-PAGEにかけたのち、蛋白をPVDF膜にプロットし、ブロッキングの後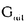 /はAS/7ポリクローナル抗体(NEN),pRbはG3-245モノクローナル抗体(Pharmingen), /はAS/7ポリクローナル抗体(NEN),pRbはG3-245モノクローナル抗体(Pharmingen),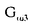 はEC/2(NEN), はEC/2(NEN),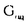 / /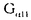 はQL(NEN), はQL(NEN),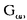 は抗 は抗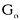 抗体(UBI)で確認した。 抗体(UBI)で確認した。 結果 gip2によるc-fosプロモーターの活性化:gip2はCHO細胞においてc-fosプロモーターCAT活性を時間依存性かつ導入したcDNAの容量依存性に活性化した。この活性は癌遺伝子Ha-rasによる活性化の60%であり、有意義な作用と考えられる。gip2の発現は導入したcDNAの容量依存性でり、このcfos転写活性はgip1,gip3,gop,gzpでは見られず、gip2特異的であった。gip2はc-mycプロモーターは活性化せず、c-fosプロモーター上のSRE,CRE,TREの中ではSRE-CATが有意に活性化された。更にSRE部位を欠失したc-fosプロモーターCATを作成し、gip2とCHO細胞に共導入した所、この変異プロモーターは高い基礎活性があり、gip2はこのSRE欠失プロモーターを全く活性化せず、強くcDNA容量依存性に抑制した。以上からgip2はSREを介してc-fosプロモーターを活性化していることが確認された。 gip2によるc-fosプロモーターのサイレンサー部位への作用:この抑制作用の結果から、gip2はc-fosプロモーター上のサイレンサー部位にも作用している事が示唆された。c-fosプロモーターには唯一pRbにより活性化されるサイレンサー(RCE)がある。gip2はRCE-CAT活性をcDNA容量依存性に強く抑制した。又、gip2とpRbの共発現は各々の単独発現より強くRCE-CAT活性を抑制した。更に、RCE部位欠失c-fosプロモーターCATを作成した所、gip2,pRbは単独発現、共発現ともにCAT活性を抑制せず、gip2は反対にCAT活性を促進した。以上よりgip2はc-fosプロモーターをRCEを介して抑制している事が確認された。次に、このgip2によるRCE-CAT活性の抑制が、pRbを介するものか確認するために、pRbの欠失したY79細胞を用いて解析した所、pRbやgip2の単独発現ではRCE-CATを全く抑制せず、pRbと共発現した時のみRCE-CATは有意に抑制された。これはgip2がpRb依存性にRCE活性を刺激している事を証明し、且つgip2がpRbを活性化していることも示唆した。 gip2のpRb及びRbプロモーターへの作用:pRbはそのセリンリン酸化状態で活性制御されている事から、gip2によるリン酸化状態への作用をpRbのウエスタンブロットで確認した。gip2はcDNA容量依存性に低リン酸化型(活性化型)pRbの発現量を増加させた。このgip2によるpRbの発現増加が転写レベルで行われているかの確認のため、gip2によるRbプロモーターCAT活性への作用を解析した所、cDNA容量依存性にCAT活性は促進した。以上よりgip2はpRbを量的且つ質的に活性化していることが確認された。 gip2のRat-1細胞に対する作用:gip2で形質転換する細胞株として知られているRat-1細胞でgip2及びpRbのc-fosプロモーターCAT又はRCE-CATに対する作用を解析した所、gip2は有意にc-fosプロモーターCAT活性を促進し、これはpRbの共発現により抑制されなかった。RCE-CATはY79細胞株の時と同様にgip2,pRbの単独発現では抑制されず、共発現でも殆ど変化が無かった。以上よりRat-1細胞ではgip2及びpRbによるc-fosの負の転写活性制御には抵抗性であった。 考察 本研究では癌遺伝子gip2が細胞内情報伝達のターゲットとしてc-fosの転写活性を促進することを示した。この作用は他の活性化型G蛋白と比してgip2特異的であり、これはは細胞増殖や癌化に関与しているのに対し、は寧ろ細胞内輸送や細胞外分泌に関与しているという最近の報告とも一致する。 次に重要な発見としてはgip2がc-fosプロモーターをSREを介して正に制御し、RCEを介して負に制御している事がある。SREを介してc-fosプロモーターを活性化するものにはHa-ras等があるが、gip2の例ではc-fosプロモーターを正負2方向に制御するためそのバランスで出力が決定されるものと思われる。pRbの大量発現によりgip2によるc-fosプロモーター活性が抑制性に変化することから、このGi2-c-fosシグナルは発生の段階で発現量が大きく変化するpRbの発生学的機能の標的である可能性がある。 gip2で形質転換されない細胞株は数多くあり、gip2型の点変異が認められるのは限られた内分泌腫瘍にしか無いことから、gip2には正負両方性の形質転換能があると考えられる。本研究では実際にgip2にはpRbを介する抑制性の情報伝達が存在しており、pRbはgip2による癌化において重要な因子である可能性がある。この考えはgip2で形質転換されるRat-1細胞においてはpRBによるRCE刺激に抵抗性があり、なお且つgip2はc-fos転写を刺激する結果からも支持される。 結論 本研究ではgip2はc-fos転写活性を促進した。gip2はc-fosプロモーター上のSREを介して正に制御し、RCEを刺激して負に制御しており、このgip2のRCEに対する作用はpRbを介するものであった。gip2はpRbの発現の促進と、pRbを低リン酸化状態(活性化型)にする事の両方でpRbを活性化していた。gip2により形質転換することが知られているRat-1細胞株では、このgip2によるpRbを介した負の転写制御が欠失していた。以上よりgip2による形質転換においてはgip2によるpRbの活性制御が重要であると結論した。 |