神経組織に豊富に存在するガングリオシドは、神経突起伸展やシナプスの形成期に著しく増大し、その組成を変化させることから、神経系の発達に重要な役割を果たしていると考えられる。神経細胞の分化や軸索伸展などに対するガングリオシドの修飾効果は、培養神経細胞を用いた多くの研究で明らかにされている。また、糖蛋白質や糖脂質上のフコース含有糖鎖は、リンパ球遊走や細胞接着との関連で最近注目されている糖鎖である。本研究では、フコース含有糖鎖の中でも、血液型抗原として古くから知られていたH抗原(Fuc 1,2Gal 1,2Gal )に注目した。 )に注目した。 神経系には、従来H抗原はほとんど存在しないと思われていたが、Ulex Europaeus agglutinin(UEA)-1 lectinや抗fucosyl GM1抗体を用いた検討により、知覚系を中心にH抗原が存在することがわかった。UEA-1 lectinの認識する2型H抗原(Fuc1,2Gal1,4GlcNAc)は、ヒトおよびウサギの脊髄後角や後根神経節神経細胞の一部に発現が認められた。後根神経節神経細胞は、末梢からの感覚刺激を中枢へ伝える1次知覚ニューロンであり、脊髄後角は知覚伝導路の経路にあたる。3型H抗原(Fuc1,2 Gal1,3GalNAc)を有するガングリオシドであるfucosyl GM1に対する抗体でも、ヒトおよびウサギの脊髄後角・後根神経節神経細胞の一部と、それを取り囲む衛星細胞が染色された。それに加えてウサギでは、脊髄前角の大型神経細胞や、後根神経を走る小径線維の軸索およびミエリンも染色された。更に、ウサギの後根神経節における、これらのH抗原の発達段階による変化をみたところ、fucosyl GM1は胎生25日頃から出現するのに対し、2型H抗原はその頃には存在せず、生後発現が見られるようになった。知覚系を含めて中枢神経系で軸索伸長やシナプス形成の盛んな出生前後にH抗原の発現が大きく変化すること、知覚神経細胞の種類によってH抗原発現が異なることは、H抗原発現と知覚神経細胞の分化や機能との関連性を示唆していると思われる。H抗原の生合成の制御機構や神経細胞での生理的意義を解明するためには、H抗原の生合成を司る1,2フコース転移酵素遺伝子のクローニングが必須と考えられた。 1,2フコース転移酵素遺伝子には、ヒトでの遺伝学的研究から少なくとも2種類の遺伝子が想定され、H遺伝子およびSe遺伝子と呼ばれていた。ヒトH遺伝子は1990年にクローニングされ、365個のアミノ酸をコードしており、他の糖転移酵素と同様にC端側をゴルジ腔に突きだした2型糖蛋白質であることが知られていたが、ヒトSe遺伝子はクローニングされていなかった。そこで本研究ではまず、ウサギの1,2フコース転移酵素遺伝子のクローニングを行った。そして、得られた1,2フコース転移酵素遺伝子を株化神経細胞に安定発現させ、細胞機能の変化を観察することによって、H抗原と神経細胞機能との関係について考察した。 ヒトH遺伝子の塩基配列を基に作製したプライマーを用いて、ウサギDNAを鋳型としてPCRをおこない、得られたDNA断片をプローブとしてウサギゲノミックDNAライブラリーをスクリーニングした。その結果、2種類のウサギ1,2フコース転移酵素遺伝子(RFT-I,RFT-II)をクローニングすることができた。RFT-IおよびRFT-IIは、ヒトH遺伝子や他の糖転移酵素遺伝子同様に2型蛋白質をコードしていると考えられた。両者とも、細胞外領域と思われるC端側に3カ所のN結合型糖鎖付加部位をもち、うち2カ所はヒトH遺伝子も含めて相同な部位であった。RFT-I,RFT-IIおよびヒトH遺伝子のアミノ酸配列の比較から、細胞外領域のグリシン(RFT-Iの79番アミノ酸、RFT-IIの71番アミノ酸)からアラニン(RFT-Iの354番アミノ酸、RFT-IIの344番アミノ酸)までは非常にホモロジーが高く、酵素活性に重要な領域であろうと推測された。RFT-IとヒトH遺伝子のアミノ酸配列の相同性は80%にのぼり、RFT-IIとヒトH遺伝子間の55%に比べて極めて高かった。しかも、RFT-IとヒトH遺伝子の相同なアミノ酸配列は、活性領域以外の全領域において認められたことから、RFT-IがウサギにおけるヒトH遺伝子の相同遺伝子であることが強く示唆された。RFT-Iは成獣ウサギの大脳・小脳に発現しており、また、脳での発現は胎児期28日目より検出できることがわかった。RFT-IIの発現は唾液腺と乳腺に認められ、胎児期の臓器には検出できなかった。 RFT-IおよびRFT-II遺伝子を発現ベクターに組み込み、COS-7細胞に一過性発現させて得た酵素標品を用いて、酵素学的検討をおこなったところ、両者とも1,2フコース転移酵素活性を有することがわかった。RFT-Iの1,2フコース転移酵素活性は、phenyl--Galあるいは1型・2型・3型糖鎖に対してほぼ同程度のKm値をとり、ヒトH酵素のそれに類似した結果であった。また、RFT-Iは糖蛋白質も糖脂質もよい基質とすることも示された。これらの結果は、RFT-Iが非還元末端の-Galのみを認識して、フコース付加をおこなっていると解釈できる。一方、RFT-IIの基質親和性は、phenyl--D-Galや2型糖鎖に比べて、1型・3型糖鎖に対して明らかに高いものであり、これまでのヒトSe酵素での報告と概ね一致していた。このことから、RFT-IIは酵素学的にSe型の1,2フコース転移酵素であると考えられた。(その後、ウサギには第3の1,2フコース転移酵素(RFT-III)遺伝子が存在し、これがヒトのSe遺伝子に対応することがわかった。) 糖鎖と細胞機能との関係を調べていくとき、遺伝子導入を容易に行える株化細胞を用いた研究が、第一段階として有用である。糖鎖骨格によって親和性の異なる2つのウサギ1,2フコース転移酵素遺伝子、RFT-I・RFT-IIを得たので、これらの遺伝子をマウス神経芽細胞腫株Neuro2a細胞に導入し、糖鎖の修飾による変化を観察した。RFT-IおよびRFT-IIをpRc-CMV発現ベクターに組み込み、Neuro2a細胞に導入後G-418存在下で培養して、遺伝子を発現している細胞をクローニングした。最初に、細胞表面の糖鎖を分析したところ、RFT-Iを発現しているNeuro2a細胞(N2A-RFT-I)ではUEA-1 lectin染色陽性(2型H抗原)・fucosyl GM1陽性(3型H抗原)だったのに対し、RFT-IIを発現しているNeuro2a細胞(N2A-RFT-II)では両者ともにほとんど検出できなかった。 Neuro2a細胞は、1 MのGM1を添加した無血清条件で培養することにより、神経突起を伸ばして分化することが知られている。Neuro2aおよびそれ由来の細胞を同条件で培養したところ、Neuro2a細胞やN2A-RFT-II細胞では1、2本の長い神経突起を伸ばした。この神経突起はリン酸化ニューロフィラメント陽性だったことから、軸索類似の性質をもつと考えられた。一方、N2A-RFT-I細胞では数本の短い神経突起のみを伸ばし、これらの神経突起は脱リン酸化ニューロフィラメント陽性であることから、樹状突起様のものと考えられた。 MのGM1を添加した無血清条件で培養することにより、神経突起を伸ばして分化することが知られている。Neuro2aおよびそれ由来の細胞を同条件で培養したところ、Neuro2a細胞やN2A-RFT-II細胞では1、2本の長い神経突起を伸ばした。この神経突起はリン酸化ニューロフィラメント陽性だったことから、軸索類似の性質をもつと考えられた。一方、N2A-RFT-I細胞では数本の短い神経突起のみを伸ばし、これらの神経突起は脱リン酸化ニューロフィラメント陽性であることから、樹状突起様のものと考えられた。 N2A-RFT-I細胞とNeuro2a細胞やN2A-RFT-II細胞との違いは、N2A-RFT-I細胞では2型H抗原・3型H抗原が発現していることであった。2型H抗原は主に糖蛋白質のN結合型糖鎖上に表現される。従って、N結合型糖鎖のプロセッシングを阻害するswainsoninを添加して培養することで2型H抗原の発現も抑制されると考えられる。実際、swainsonin存在下で培養することによってN2A-RFT-I細胞のUEA-1 lectinに対する染色性は著明に低下するものの、無血清条件での樹状突起様神経突起の形成は変化しなかった。次に、ガングリオシド生合成を抑制するD-threo-PDMPを添加して培養すれば、fucosyl GM1の発現も結果的に抑制されると考えられる。D-threo-PDMP存在下ではN2A-RFT-I細胞におけるfucosyl GM1の発現はほとんど消失し、同時に、無血清培養条件でリン酸化ニューロフィラメント陽性の長い神経突起を伸ばすようになった。更に、このような軸索様神経突起の伸展は、D-threo-PDMPと-L-fucosidase共存在下で培養することによって一層著明になった。D-threo-PDMPを添加した培養条件では、ガングリオシド全体の生合成が抑制されているので、結果の解釈は慎重を要するものの、fucosyl GM1発現の有無が神経突起の性質を変化させていると考えられた。-L-fucosidase処理をした場合、fucosyl GM1が分解されてGM1が生成するので、GM1の添加と同効果で軸索伸展を促進する結果になった解釈できる。以上の実験から、Neuro2a細胞表面のガングリオシドの修飾と、神経突起伸展の表現型の変化とを、シェーマにまとめて下図に示す。 細胞表面のガングリオシドの修飾が、最終的に神経突起伸展という形態変化につながるまでの情報伝達経路は、これまでのところ全くわかっていない。様々な形で糖鎖を変異させた細胞を比較し、セカンドメッセンジャー系や様々なカイネース系などを解析することが、今後必要であろう。  図1:1,2フコース転移酵素遺伝子発現による糖鎖の修飾と、神経突起伸展の表現系との関係。 図1:1,2フコース転移酵素遺伝子発現による糖鎖の修飾と、神経突起伸展の表現系との関係。 | 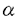 1,2-フコース転移酵素遺伝子の発現による神経芽細胞腫Neuro2a細胞の軸索伸長抑制
1,2-フコース転移酵素遺伝子の発現による神経芽細胞腫Neuro2a細胞の軸索伸長抑制