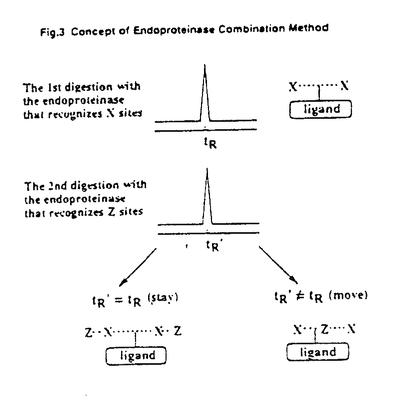
本論文は核内レチノイン酸受容体のリガンド結合ドメインについて、その三次構造に関する情報を有機化学的な手法により、簡便に得るべく展開した研究を記述している。 レチノイン酸はビタミンAの活性本体であり、哺乳動物の生命維持・正常な成長などに必須な因子である。その細胞分化・増殖に及ぼす決定的な生物作用から、医薬応用の観点からも注目され、多くの誘導体(レチノイド)がデザイン・合成されている。レチノイドの作用機構は、核内レチノイン酸受容体を介した特異的な遺伝子の発現制御にある。核内レチノイン酸受容体にはいくつかのサブタイプが存在し、類縁受容体として多くのステロイドやチロキシンホルモンなどの受容体がある。新規レチノイン酸誘導体、特にサブタイプ選択的なレチノイドの開発に際しては、核内レチノイン酸受容体のリガンド結合ドメインにおいて直接リガンドと接するペプチド鎖を特定することが重要な課題となる。 そこで佐々木は本課題を解決するために、(1)核内レチノイン酸受容体リガンド結合部位に対して特異的な光親和性標識化合物を複数デザイン・創製し、(2)これらを用いて光標識した部位を簡便に特定するために、エンドプロテアーゼコンビネーション法と呼ぶ一般的な手法を開発し、(3)本方法により、核内レチノイン酸受容体においてリガンドと直接相互作用すると目される三つの部位をアミノ酸配列のレベルで特定した。 光親和性標識剤としては、既知の強力な合成レチノイドのカルボキシル基末端近傍にスペーサーを介してアジドダンシル基を導入したものと、類似の分子骨格の疎水性頭部に直接光反応性のトリフルオロメチルジアジリジン基を導入したものをデザイン・合成している。特に前者の創製に至る過程では、核内レチノイン酸受容体を蛍光強度の変化の測定のみで簡便に検出し得る蛍光性レチノイドブローブの開発にも成功している。 創製した光親和性標識剤により特異的に核内レチノイン酸受容体を光標識し得ることを、リコンビナントなヒト核内レチノイン酸受容体リガンド結合ドメインタンパクを用いて確認している。 次いで佐々木は、標識タンパクの標識剤結合部位を簡便にアミノ酸配列のレベルで特定し得る、一般応用性のある新手法、エンドプロテアーゼコンビネーション法を開発した。この方法は、標識タンパクをアミノ酸残基特異的なエンドプロテアーゼ複数種を段階的に用いて消化し、各段階における標識フラグメントの異同を高速液体クロマトグラフィー分析によって確認し、その結果からエンドプロテアーゼが認識するアミノ酸に関する一次構造上の相互位置関係(コンテクスト)を決定するというものである。本方法は、一次構造の決定されているすべてのタンパクについて応用可能な一般的かつ簡便な方法である。 本論文では標識ヒト核内レチノイン受容体リガンド結合ドメインタンパクについて、四種のアミノ酸残基特異的エンドプロテアーゼを用いたエンドプロテアーゼコンビネーション法が適用され、標識部位一ヶ所につき12項目のアミノ酸コンテクストを得ている。得られた結果に合致するコンテクストを既知の核内レチノイン酸受容体一次アミノ酸配列の中に探索するという方法により、2種の光親和性標識剤について合計三ヶ所の標識部位が特定された。 本論文で佐々木が特定した上記三ヶ所の標識部位は、ヒト核内レチノイン酸受容体リガンド結合ドメインの、ヘリックス8中のアルギニン残基、ヘリックス6中の9-アミノ酸残基からなる部位、およびヘリックス12中の3-アミノ酸残基からなる部位である。これらの部位はいずれもリガンド結合ポケットを形成する部分に存在し、佐々木の方法の有効性が実証されている。 本論文による、核内レチノイン酸受容体がリガンドと直接相互作用するアミノ酸部分の特定により、受容体サブタイプ選択的なレチノイドのデザイン展開が期待できる。加えて本論文に記述された全体的なストラテジーは、その有効性の実証と一般性から、広く薬物受容体・標的分子の構造解析ならびにそれらの特徴ある新規リガンドのデザインに向けて期待できる新手法である。 よって本研究は、生物有機化学・医薬化学の進展に寄与するところ大きく、博士(薬学)の学位にふさわしいものと判断した。 |  の特異的標識を行なった。さらに、標識部位を解析するために「エンドプロテアーゼコンビネーション法」を開発・確立し、本法の応用によって標識された部位の特定を行なった。
の特異的標識を行なった。さらに、標識部位を解析するために「エンドプロテアーゼコンビネーション法」を開発・確立し、本法の応用によって標識された部位の特定を行なった。


 リガンド結合領域とall-trans-レチノイン酸との複合体のX線結晶構造解析が行なわれ、核内レセプター間で共通すると考えられるホロフォームの基本構造が示されたが、hRAR
リガンド結合領域とall-trans-レチノイン酸との複合体のX線結晶構造解析が行なわれ、核内レセプター間で共通すると考えられるホロフォームの基本構造が示されたが、hRAR