| 内容要旨 | | 陸棲軟体動物コウラナメクジ(Limax marginatus)は比較的単純な神経系を持つにもかかわらず高度な嗅覚-味覚連合学習を行うため,学習・記憶の優れたモデル系と考えられる.コウラナメクジの嗅覚-味覚連合学習には脳神経節の前脳と呼ばれる部位の関与が示唆されている.前脳は中枢神経系の大部分に相当する数(105個程度)の介在神経からなり,触角からの神経線維の最も主要な投射先である.前脳の介在神経は同期した約0.7Hzの膜電位振動を定常的に生じており,膜電位振動は嗅覚刺激や味覚刺激によって変調を受ける.学習の機構を解明するためには,膜電位振動の生成・修飾機構を含む神経回路の機能的構造を明らかにすることが重要である.しかし前脳の神経細胞は細胞体が小さく通常の細胞内記録が困難なため,これまで単一細胞レベルでの神経伝達に関する研究はほとんど行われていなかった. そこで本研究でははじめに単一の前脳ニューロンの電気的活動と形態の関連を明らかにすることにより前脳ニューロンの機能について検討した.軟体動物で広く存在して多様な働きをするグルタミン酸の単一ニューロンに対する効果を穿孔パッチ(perforated patch)法を用いて調べた.次にに回路全体で同期した膜電位振動に対するグルタミン酸の修飾効果を単一細胞に対する効果と比較し,電気生理学的方法では測定困難な神経突起部位に対する効果を光学測定によって検討した. [結果]1.前脳ニューロンの形態 コウラナメクジの脳神経節を単離し,前脳の細胞体層部分をdesheathして正立顕微鏡下で単一神経細胞からナイスタチン穿孔パッチ法により記録を行った.電流固定モードで記録される自発的な活動から,前脳神経はbursting neuronとnonbursting neuronに分類されることが確認された(図 1A,B). bursting neuronは周期的に脱分極入力を受けてバースト状に発火し,nonbursting neuronはバーストを生じず周期的に強い過分極を受ける.これらは前脳内で混在していたが,bursting neuronはnonbursting neuronに比べ細胞体が大きく,長い形態のものが多い特徴を持つことから、両者が形態学的にも区別されることが明らかになった(図1C).次にsulforhodamine Bの細胞内注入によってそれぞれの細胞の形態を調べた.その結果,bursting neuronと考えられる細胞体の長径が9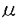 m以上の細胞では,dorsal方向から見ると突起をcell massとterminal massの境界付近へ伸ばしながら数本に分岐しているのが観察されたが,internal massへは突起をのばしていなかった.またこれをposterior方向から見るとsagittal方向に数本の突起を伸ばしていた.一方,nonbursting neuronと考えられる長径が8m以下の細胞では,dorsal方向から見るとterminal massを通過してinternal massへ到達しているのが観察された. m以上の細胞では,dorsal方向から見ると突起をcell massとterminal massの境界付近へ伸ばしながら数本に分岐しているのが観察されたが,internal massへは突起をのばしていなかった.またこれをposterior方向から見るとsagittal方向に数本の突起を伸ばしていた.一方,nonbursting neuronと考えられる長径が8m以下の細胞では,dorsal方向から見るとterminal massを通過してinternal massへ到達しているのが観察された.  図1 (A),(B)前脳ニューロンの自発的活動の電流固定モードによる記録.(A)bursting neuron,(B)nonbursting neuron,(C)bursting neuronとnonbursting neuronの細胞体の大きさ(長径)の分布. 図1 (A),(B)前脳ニューロンの自発的活動の電流固定モードによる記録.(A)bursting neuron,(B)nonbursting neuron,(C)bursting neuronとnonbursting neuronの細胞体の大きさ(長径)の分布. 図2 前脳ニューロンに対するグルタミン酸の効果.(A)nonbursting neuron,(B)bursting neuron.2.前脳ニューロンに対するグルタミン酸の局所投与の効果 図2 前脳ニューロンに対するグルタミン酸の効果.(A)nonbursting neuron,(B)bursting neuron.2.前脳ニューロンに対するグルタミン酸の局所投与の効果 単一の前脳ニューロンから電位固定モードで記録を行い,細胞体近傍にマイクロインジェクターを用いてL-グルタミン酸(100-500M)を投与すると,2種類のニューロンのそれぞれに特徴的なグルタミン酸応答が生じた(図2).nonbursting neuronでは持続的な外向き電流(反転電位約-90mV)がみられ,この電流は電流-電圧関係と外液イオン組成に対する依存性からK+電流と考えられた.一方bursting neuronでは主として一過的な電流(反転電位約-50mV)が顕著にみられ,この電流は電流-電圧関係と外液イオン組成に対する依存性からCl-電流と考えられた.bursting neuronではこれに加えて小さい持続的なK+電流と考えられる外向き電流成分も生じた.nonbursting neuronにおける自発的な外向き電流もグルタミン酸によって生じるK+電流と同様の電流-電圧関係及びイオン選択性を示し,またこれらの電流は非相加的であったことから,自発的な電流はグルタミン酸を伝達物質とするシナプス電流の可能性が高いと考えられた. グルタミン酸レセプターアゴニストとして知られている薬物の中で,キスカル酸は選択的に持続的なK+電流応答を生じ,イボテン酸は選択的に一過的なCl-電流応答を生じた.これらの薬物はグルタミン酸と同濃度でほぼ同程度の効果を示した.L-AP4,カイニン酸,trans-ACPD,AMPAはいずれも1mMで電流応答をほとんど生じなかった.またグルタミン酸レセプターアンタゴニストであるCNQX(500M)は2種類のグルタミン酸応答のいずれも阻害しなかった. 3.薬物の灌流投与による膜電位振動の変化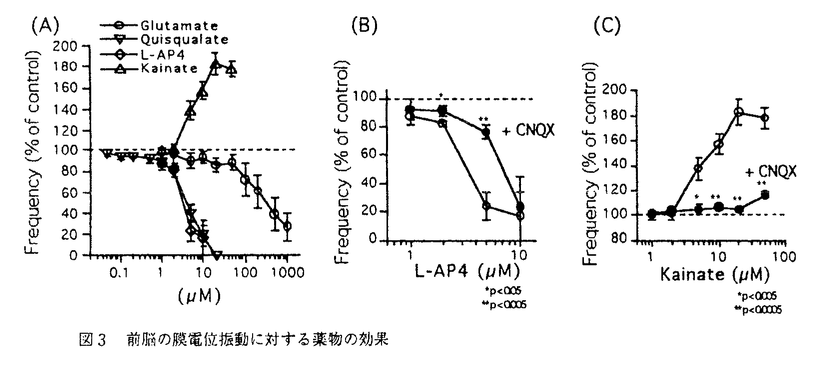 図3 前脳の膜電位振動に対する薬物の効果 図3 前脳の膜電位振動に対する薬物の効果 グルタミン酸の膜電位振動に対する修飾効果を,薬物の灌流投与により検討した.膜電位振動は前脳の表面から細胞外電極を用いて記録される局所場電位によりモニターした.グルタミン酸は比較的高濃度で膜電位振動の振動数を減少させた.キスカル酸,L-AP4はグルタミン酸より約100倍低濃度で膜電位振動の振動数を減少させた.カイニン酸は逆に膜電位振動の振動数を増大させた(図3A).L-AP4とカイニン酸の効果はCNQX(50M)によって抑制された(図3B,C)が,自発的な膜電位振動の振動数にはCNQXは効果がなかった. 4.内在性の神経伝達物質による膜電位振動の修飾 触角神経束を吸引電極を用いて単発電気刺激すると,多くの試料で10s程度にわたって膜電位振動の振動数が減少した.CNQX(500M)をあらかじめ灌流しておくとこの振動数の減少は阻害された.一方電気刺激により振動数が増大がみられる場合もあったが,これはCNQXでは阻害されなかった.このことから,前脳に対する触角神経束からの入力による膜電位振動の修飾のうち,振動数の減少はグルタミン酸によって担われていることが示唆された. 5.薬物による膜電位変化のイメージング 以上の結果から単一細胞でみられる電流応答と膜電位振動に対する効果が薬物によって異なることが示された(表1).この結果は膜電位振動の修飾に前脳ニューロンの細胞体以外の部位が関わっていることを示唆している.そこで薬物の作用部位を明らかにする目的で薬物による膜電位変化のイメージングを行った.膜電位変化の画像は,神経節を電位感受性蛍光色素Di-4-ANEPPSで染色し,蛍光顕微鏡とフォトダイオードカメラシステム(HR-Deltaron1700)を用いて,初期画像に対する蛍光強度の差分として取得した.その結果,振動数を減少する薬物(グルタミン酸,キスカル酸,L-AP4)は前脳の平均膜電位を過分極し、振動数を増大する薬物(カイニン酸)は平均膜電位を脱分極するのが観察された.このことから、電位振動の修飾には前脳ニューロンの膜電位変化が関わっていると考えられた.  表1 前脳に対する薬物の効果[まとめ] 表1 前脳に対する薬物の効果[まとめ] 2種類の前脳ニューロンの形態とグルタミン酸応答が明らかになり,これらのニューロンの機能の違いが示唆された.グルタミン酸関連薬物には膜電位振動に対する選択的な効果もあり,前脳の情報処理過程における関与が示唆された. |