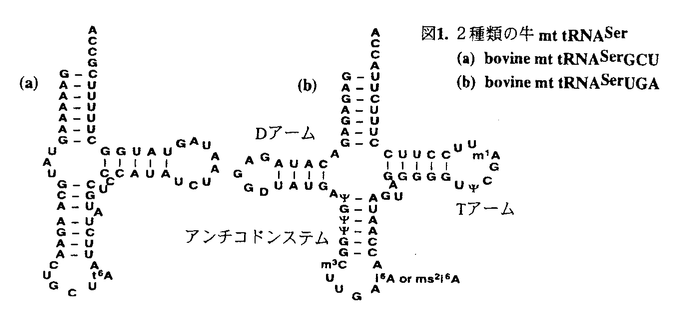
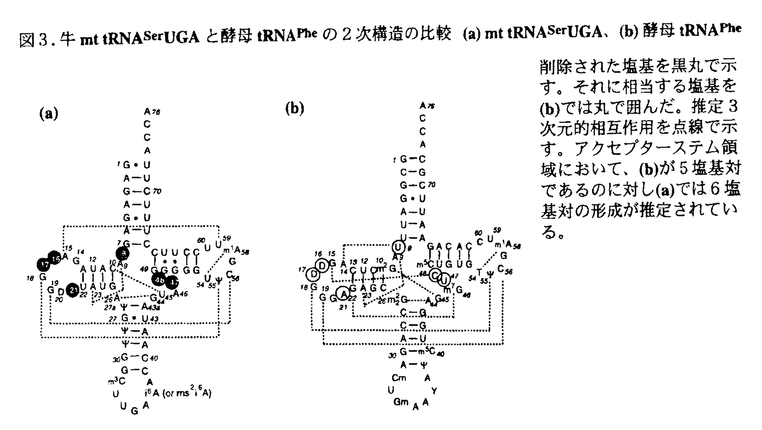
動物ミトコンドリアのtRNAは細菌や細胞質の通常のtRNAとは著しく異なった構造を持つことが主としてミトコンドリアDNAの解析から推定されてきた。これらのtRNAの中には通常のDループ・Tループ間相互作用を欠いているものばかりでなくDアームやTアームが欠落しているものの存在も知られている。これまでRNAレベルでの生化学的な実験によって、これらのtRNAの1次および2次構造解析が行われ、またこのような異常tRNAが実際に生体内で機能していることも明らかにされつつある。これらの異常なミトコンドリアtRNAは、tRNAの機能構造を根本的に理解する上に有力な素材として存在していたが、これまで試料調製の困難さからそれらの機能構造の研究は殆ど未開の分野として残されていた。本研究はこの課題に果敢に挑戦したものである。 哺乳動物のミトコンドリアには、UCNコドン(N;A,T,G,C)とAGYコドン(Y;C,T)に対応する2種類のセリンtRNA(tRNASerUGAとtRNASerGCU)が存在しているが、前者は通常のクローバーリーフ型に近い2次構造をもつのに対して後者はDアームを欠くという全く異なった2次構造を有している。それにも拘わらず、両tRNAは同一のミトコンドリアのアミノアシルtRNA合成酵素に認識される。この認識機構を知るためにも、また高次構造を明らかにするためにも、これらの2種類のtRNAの立体構造を決定する必要がある。本論文ではイミノプロトン領域のNMRを測定することにより、生化学的な実験から推定されたこれら両tRNASerの2次構造および3次元相互作用を検証し、さらにコンピューターモデリングからその機能構造を検討した。これらを第1部とし、第2部ではこれら異種tRNAの大腸菌内での発現系の構築について述べている。 第1部の第1章はtRNASerUGAの立体構造の解析結果である。tRNASerUGAは通常のtRNAに酷似した2次構造と3次元相互作用をもつにも関わらず、これまで調べられたtRNAの中で最も低い融解温度(Tm)を示す点で、特にその立体構造に興味が持たれた。NMR測定には大量(数mg)の試料を必要とするが,動物ミトコンドリアから数mgのtRNAを調製することは大変困難であるため、試験管内でT7RNA polymeraseによる転写合成を行った。この未修飾tRNAが天然のtRNAと共通な構造を保持していることは、ヌクレアーゼや化学試薬による感受性の観察とアミノ酸受容能から確認された。この未修飾tRNAについて、1次元NOE測定及び安定同位体標識NMR法の利用によって、イミノプロトン領域のシグナル帰属がほぼ完了し、すでに生化学的実験に基づいて推定されていた、L字型の立体構造をとるというコンピューターモデルの正当性が明示された。さらにこのtRNAのマグネシウム結合部位について検討したところ、通常のtRNAと全く異なり、イミノプロトンNMRスペクトル上ではDアーム領域にあたるシグナルの化学シフトに殆ど変化が観測されなかった。コンピューターモデルでDアーム近傍の構造を調べたところ、通常のtRNAの結晶構造中でDアームに配位している2つのマグネシウムイオンがこのtRNASerUGAでは配位できなくなっていることがわかり、このことがこのtRNAの異常なTmの低さの原因であると推測された。 第2章ではDアームの欠けたtRNASerGCUのNMRスペクトルの解析結果について述べている。ここでも試料としてT7RNA polymeraseによる転写物を用いているが、このtRNAは修飾塩基としてアンチコドン3’端に隣接した23位にt6Aのみしか含まないことから、未修飾tRNAのアミノ酸受容能は天然のものとよい一致を示した。このtRNAのNMRスペクトルは特にシグナルの重複が激しく、塩基対置換体や部位特異的な安定同位体標識によってもG-C塩基対由来のイミノプロトンを帰属することが可能になったのみで、A-U塩基対由来のイミノプロトンのシグナルについては帰属することができなかった。G52のシグナル強度が非常に弱いこと、アクセプターステムとTステムをつなぐ塩基対(G52/G51)の間でNOEを観測することができなかったことから、このtRNAはDアームの欠如をTアームの構造変化によって補っているということが推測された。さらに帰属のついたシグナルについてマグネシウム滴定実験を行った結果、tRNAのコア領域にあるG-C塩基対(G11,G31)の化学シフトはマグネシウム濃度に対してほとんど変化を示さなかった。このtRNASerGCUにおいてもDアームの欠けた特異な構造をとるためにマグネシウムイオンの結合様式が通常のtRNAとは異なっていることが推測された。 第2部はこれら異常なミトコンドリアtRNAを大腸菌内で大量に発現させる系の開発について述べたものである。これまで利用してきたT7RNA polymeraseを用いた転写合成法ではその転写効率がプロモーター下流の配列に大きく依存すること、また転写産物の3’末端が均一にはならないことが問題となっていた。そこで大腸菌を用いた大量発現系の構築を試みた。大腸菌を用いた異種tRNAの発現系はこれまでほとんど報告されておらず、異常構造をもつtRNAについては全く成功してはいなかった。この原因がtRNAの安定性にあるとの予想の基に、tRNA遺伝子のアクセプターステム領域をA-U塩基対からG-C塩基対に置換して大腸菌内の発現系に導入したところ、100mLの培地から0.4mgのミトコンドリアtRNA変異体(修飾塩基は含まないが、5’端,3’端ともに均一にそろっている)の調製に成功した。このtRNA変異体はTmが天然型に比べて5℃高く、X線結晶解析の試料として利用できる可能性の高いこと、最小培地でも十分量のtRNAの発現が期待でき、NMR測定に有用な同位体標識法が利用できることなど、異種tRNAの大量調製法として有用な新しい方法を確立したと考えられる。 以上要するに、本論文は異常なミトコンドリアtRNAのNMRスペクトルを初めて測定し、その立体構造とマグネシウム結合部位について新たな知見を得、さらに大腸菌内で異種tRNAを発現する新しい方法を開発したものであり、翻訳系のメカニズムの解明とその利用に大きく貢献した。従って本論文は生体系を利用した蛋白質生産の基礎研究として工学に資するところ大である。よって本論文は博士(工学)の学位請求論文として合格と認められる。 |