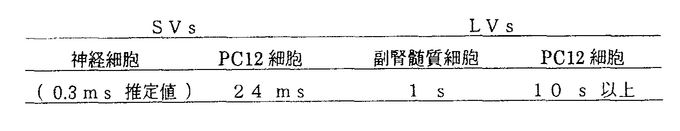
本研究は、開口放出における分泌小胞の膜融合速度を測定し定量化するために、PC12細胞および副腎髄質細胞にケイジド試薬を利用しCa2+シグナルの多様性を回避して膜容量測定法と単一小胞分泌の直接測定法を組み合わせて分泌小胞の種類を特定したうえでその膜融合時定数の直接測定を試みたものであり、下記の結果を得ている。 I.PC12細胞には速い開口放出とエンドサイトーシスおよびそれに続く遅い開口放出とエンドサイトーシスが存在し、その膜面積の大きさにおいてそれぞれが強い相関関係を持ちよくバランスしていることが示された。 この結果とこれら二種類の開口放出とエンドサイトーシスの特徴から、PC12細胞にはメカニズムの異なる二つのエンドサイトーシス機構が備わっており、この二つの機構が、速い開口放出と遅い開口放出に伴い加わった膜を識別して膜の取り込みを行なっていることが推定された。 さらにPC12細胞はsmall clear vesicles(SVs)とlarge dense-core vesicles(LVs)の二種類の分泌小胞を持ちそれぞれの小胞内に選択的にacetylcholine(ACh)とmonoamines(MAs)を含むことが既に生化学的に知られており、膜容量トレースの特徴からSVsが速い開口放出を起こし、LVsが遅い開口放出を起こしているという仮説が導かれた。 II.この仮説を証明するために、AChおよびMAsの分泌の直接測定が行われ、それぞれの時間経過を速い膜容量増大および遅い膜容量増大の時間経過との対応をとることにより確かめられた。AChの測定は、ニコチニックACh受容体を豊富に発現している培養骨格筋細胞をバイオセンサーとして用いて行い、MAsの測定は、炭素繊維電極を用いて行われた。 1. AChの量子的放出頻度の時間分布図と膜容量変化の時間微分とを比較した結果AChの量子的放出は、速い膜容量の増加に時間的によく対応しており、 (1) PC12細胞からのAChの量子的放出がSVsの開口放出により起きる。 (2) SVsの開口放出は、速い膜容量増大に伴って起きる。 (3) PC12細胞のSVsの膜融合時定数が約24msである。 ことが示された。 2.MAsの量子的放出頻度の時間分布図と膜容量変化の時間微分とを比較した結果MAsの量子的放出は、遅い膜容量の増加に時間的にほぼ対応しており、 (1) PC12細胞からのMAsの量子的放出が、LVsの開口放出により起きる。 (2) LVsの開口放出は、遅い膜容量増大に伴って起きる。 (3) PC12細胞のLVsの膜融合時定数が10s以上である。 ことが示された。 3.さらに副腎髄質細胞のLVsの開口放出によるMAs分泌に関しても、炭素繊維電極を用いて同様の実験が行われた。その結果、副腎髄質細胞のLVsの開口放出の時間経過は、PC12細胞のそれより著しく速く、その膜融合時定数は、約1sであることが示された。この様に分泌小胞の種類が同じでも細胞の種類が異なると膜融合時定数が大きく異なることが示された。 以上、本論文はPC12細胞という同一細胞内の二種類の異なる分泌小胞を同定したうえで、それぞれの分泌小胞の膜融合時定数の直接測定に初めて成功したものであり、この膜融合時定数は分泌小胞の種類および細胞の種類により1000倍を超える違いがあることを明らかにしたものである。この様に本研究はこれまで調べられていなかった膜融合機構の速度的多様性を直接定量化して示したものであり、今後の開口放出機構の解明に重要な貢献をなすものと考えられ、学位の授与に値するものと考えられる。 |