| 内容要旨 | | 緒言 1924年両生類胚を用いたSpemannとMangoldの報告以来,脊椎動物胚の神経系の発生には中軸中胚葉が未分化外胚葉に働きかける神経誘導が必須な過程であると考えられているが,その分子的機構は未解明な部分が多い. 脊椎動物胚の原型とも言われている原索動物ホヤ胚は1905年のConklinの古典的な研究以来自律性分化の典型例と考えられてきたが,1946年ReverberiとMingantiにより神経系の発生には予定脊索領域を含む割球が必要である事が報告された.さらに1988年岡戸と高橋により,ホヤ8細胞胚より単離しサイトカラシンB(CB)で分裂抑止した動物半球頭側の予定神経割球a4-2は単独では表皮型細胞に分化するが,正常胚の64細胞期に相当する時期に植物半球頭側の予定脊索割球A4-1と接着して細胞間作用を受けると神経型細胞に分化しNa活動電位を示す事が報告された.この高橋等の系は極めて単純な2細胞系であるが脊椎動物胚における神経誘導現象に相同の分化過程を示しており,神経誘導の研究に極めて有用と考えられる. Spemann以来神経誘導の研究は主に両生類胚を用いて行われ,多くの研究者がその誘導信号の実体解明に努力したが,1990年代に入ってようやくLamb等によりnogginが,見学と岡本により塩基性線維芽細胞増殖因子(bFGF)がその候補として挙げられるに至った.本研究ではbFGFが原索動物のホヤ予定外胚葉割球においても神経誘導を起こす事を示すと同時に,単純な1あるいは2細胞系での誘導過程の解析によりbFGFの作用様式が内因性の予定中胚葉割球と同じである事を明らかにし、bFGFが脊椎動物胚に普遍的な誘導因子である可能性を示唆した. 実験方法 実験にはアカボヤHalocynthia aurantium初期胚を用いた.8細胞期にコリオン膜を除去し,以後は細胞分裂を抑止するためにCB2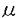 g/mlを加えた海水中で飼育した.予定神経領域および予定表皮領域を持つ動物半球頭側割球a4-2を8細胞胚から単離し,分裂抑止下に、単独培養するか,予定脊索領域を含む植物半球頭側割球A4-1と接着培養した(a-A pair).単独培養の一部にはbFGFあるいはセリンプロテアーゼの一種subtilisinによる神経誘導を施した.神経細胞への分化は,神経特異的なイオン電流の検出により確認した.また,電流記録から神経細胞と判定された割球を集め,mRNAを抽出してRT-PCR法によりホヤ神経細胞に特異的なNaチャネル遺伝子TuNaIの転写を確認する実験も行った. g/mlを加えた海水中で飼育した.予定神経領域および予定表皮領域を持つ動物半球頭側割球a4-2を8細胞胚から単離し,分裂抑止下に、単独培養するか,予定脊索領域を含む植物半球頭側割球A4-1と接着培養した(a-A pair).単独培養の一部にはbFGFあるいはセリンプロテアーゼの一種subtilisinによる神経誘導を施した.神経細胞への分化は,神経特異的なイオン電流の検出により確認した.また,電流記録から神経細胞と判定された割球を集め,mRNAを抽出してRT-PCR法によりホヤ神経細胞に特異的なNaチャネル遺伝子TuNaIの転写を確認する実験も行った. 結果 1.8細胞胚から分離した予定神経領域および予定表皮領域を持つa4-2割球を,CBを用いて分裂抑止して培養したところ従来の報告通り表皮型細胞に分化したが,bFGFを作用させると神経特異性イオンチャネルを持つ神経型細胞に分化した.Subtilisinを作用させた場合も,予定中胚葉領域を持つA4-1割球と接触培養した場合も従来の報告通り神経型細胞に分化した. 2.他の増殖因子のうちTGF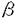 1,TGF2,activin A,EGF,NGFは効果が認められず,酸性線維芽細胞増殖因子(aFGF)のみが部分的な神経誘導活性を示した. 1,TGF2,activin A,EGF,NGFは効果が認められず,酸性線維芽細胞増殖因子(aFGF)のみが部分的な神経誘導活性を示した. 3.bFGFによる神経誘導は,RT-PCR法により,ホヤ神経細胞に特異的なNaチャネル遺伝子TuNa Iの転写としても証明された. 4.bFGFは1ng/mlで既に10%以上の割球を神経分化させ,100ng/ml以上では効果はほぼ飽和し,90%の割球を神経分化させた.濃度依存曲線はEC508ng/mlのミカエリス・メンテン曲線でほぼ近似できた. 5.神経誘導効果を最大限に発揮するためには,bFGFは受精後8時間目から14時間目まで持続的に作用する事が必要であり,A4-1割球による神経誘導が完成するには,受精後8時間目から14時間目まで持続的に接触している必要がある点と一致した. 6.bFGFによる神経誘導の結果内向き整流性K電流の増加が受精後20時間以後より減少に転じ,subtilisinあるいはA4-1割球との接触による神経誘導における従来の報告と一致した. 7.Subtilisin処理したa4-2割球にチロシンキナーゼ抑制剤LC6を作用させたところ表皮分化が回復し,bFGF受容体のチロシンキナーゼ活性化が,bFGF・subtilisin・A4-1割球との接触という3通りの神経誘導に共通の経路である可能性が示唆された. 考察と結語 本研究は,両生類における神経誘導因子研究の示唆より,脊椎動物の原型と考えられる原索動物においてbFGFの効果を解析し,bFGFの神経誘導作用を原索動物においても確認した.この結果,従来両生類を用いた複雑な多細胞系で研究されてきた神経誘導機構を,単純な原索動物の単一細胞系によって,脊椎動物に共通な機構として解析できる可能性を示したと言える. |
| 審査要旨 | | 本研究は,脊椎動物の神経系の発生に必須な神経誘導現象における塩基性線維芽細胞増殖因子(bFGF)の役割を,原索動物初期胚を用いて検討し,以下の結果を得ている. 1.8細胞胚から分離した予定神経領域および予定表皮領域を持つa4-2割球を,サイトカラシンB(CB)を用いて分裂抑止して培養したところ従来の報告通り表皮型細胞に分化したが,bFGFを作用させると神経特異性イオンチャネルを持つ神経型細胞に分化した.プロテアーゼの一種subtilisinを作用させた場合も,予定中胚葉領域を持つA4-1割球と接触培養した場合も,従来の報告通り神経型細胞に分化した. 2.他の増殖因子のうちTGF1,TGF2,activin A,EGF,NGFは神経誘導効果が認められず,酸性線維芽細胞増殖因子(aFGF)のみ部分的な活性を示した. 3.bFGFによる神経誘導は,RT-PCR法により,ホヤ神経細胞に特異的なNaチャネル遺伝子TuNa Iの転写としても証明された. 4.bFGFは1ng/mlで既に10%以上の割球を神経分化させ,100ng/ml以上では効果はほぼ飽和し,90%の割球を神経分化させた.濃度依存曲線はEC508ng/mlのミカエリス・メンテン曲線でほぼ近似できた. 5.神経誘導効果を最大限に発揮するためには,bFGFは受精後8時間目から14時間目まで持続的に作用する必要があり,A4-1割球による神経誘導が完成するには,受精後8時間目から14時間目まで持続的に接触している必要がある点と一致した. 6.bFGFによる神経誘導の結果,内向き整流性K電流の増加が受精後20時間以後より減少に転じた.この点もsubtilisinあるいはA4-1割球との接触による神経誘導における従来の報告と一致した. 7.Subtilisin処理したa4-2割球にチロシンキナーゼ抑制剤LC6を作用させたところ表皮分化が回復し,bFGF受容体のチロシンキナーゼ活性化が,bFGF・subtilisin・A4-1割球との接触という3通りの神経誘導に共通の経路である可能性が示唆された. 以上本論文は,脊椎動物の原型と考えられる原索動物においてbFGFの神経誘導効果を確認し,従来両生類を用いた複雑な多細胞系で研究されてきた神経誘導機構を,単純な原索動物の単一細胞系によって,脊椎動物に共通な機構として解析できる可能性を示したと言え,学位の授与に値するものと考えられる. |