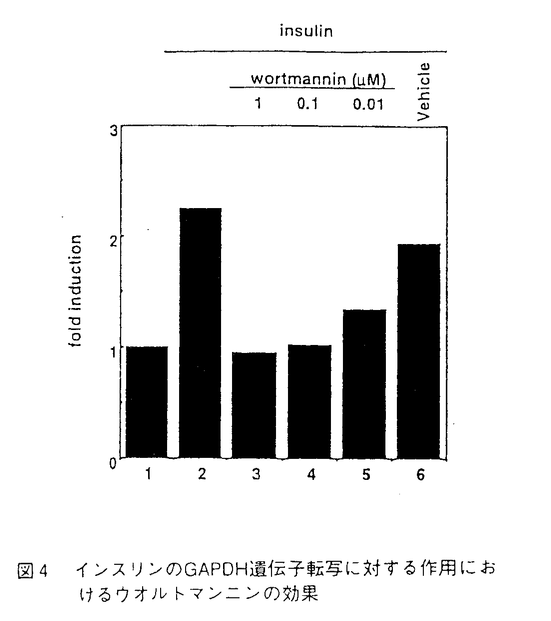
学位申請者は学位申請論文に示された研究において解糖系酵素の遺伝子発現機構について解析している。解糖系酵素のうちグリセルアルデヒド3-リン酸デヒドロゲナーゼ(以下、GAPDHと略す)遺伝子を解析の対象として選んでいるが、それはこの遺伝子がは多くの組織で構成的かつ強力に発現しているからである。その強力なプロモーター活性をもたらしている領域を同定するために、ヒトGAPDH遺伝子の-489bpから-21bpの部分に存在しているプロモーター領域をクローン化し解析している。-489bpから転写開始点に向かって順次欠失させたプロモーターを作成しCAT assayにより分析した結果、細胞種によらず-181bpから-144bpの領域を欠失させると大幅な転写活性の低下が見られることを明らかにしている。この領域を転写開始点より遠い方から順に、10個の連続したAよりなる領域 (A-stretchサイト)、GCに富んだ領域(GC-richサイトI)、AGに富んだ領域(AG-richサイト)、GCに富んだ領域(GC-richサイトII)の4サイトに便宜的にわけてそれぞれの機能を検討している。すなわち、申請者は-181bpより-144bpまでの領域に対応するDNA及び各サイトに変異を導入したDNAを合成しゲルシフト法により分析を行いA-Stretchサイトを認識して結合する因子(GAPBF1)の存在とGC-richサイトIを認識して結合する因子(GAPBF2)の存在を示した。更にA-stretchサイトの長さを順次短くしていき、GAPBF1は連続するA塩基数を8塩基にしても10塩基の場合と同じように結合するが、6塩基にすると結合活性が弱まり4塩基にすると全く結合しなくなるような因子であることを明らかにした。A-stretchサイトのシスエレメントとしての機能についても解析を行い、連続するA塩基の数を減少させると転写活性化能の減少が見られ、6塩基にするとほぼ転写活性化能が喪失することを示した。GAPBF1のDNA結合活性とA-stretchサイトの転写活性化能の塩基配列依存性は類似した傾向を示し、GAPBF1が実際に転写活性化因子として働いていることが強く示唆されている。次に、GC-richサイトIに変異を導入しても転写活性化能が減少することを明らかとしGC-richサイトIもシスエレメントとして機能していることを確認している。GAPBF2はGCに富んだ領域に結合することから典型的なGC-box結合因子であるSp1である可能性が考えられたが、申請者は抗Sp1抗体を用いた実験からGAPBF2はSp1ではないことを確認した。さらに、-144bp付近の配列を完全な形で含むように-150bpまでの領域を含むプロモーターを作りそのプロモーター活性を-143bpまでの領域を含むプロモーター活性と比較から-144bp付近にもう1つのシスエレメントの存在を推定し、この領域の配列(-149bpから-137bp)をもつ合成DNAをプローブとしてゲルシフト分析を行い、この領域に特異的に結合する因子(GAPBF3)の存在も確認した。 上述のように申請者はGAPDH遺伝子の構成的発現に必要なプロモーター主要領域とそこに結合する3種類の転写因子の存在を示した。この中で、GAPBF1の結合配列は極めてユニークなものである。なお、A-stretch配列は酵母において多くの遺伝子の構成的発現に重要であることがすでに示唆されているが、そこに結合する転写因子の介在は示されておらず、むしろA-stretch配列の特異な立体構造自体に何らかの機能が求められている。申請者の発見はこの配列を認識する蛋白質因子をつかまえている点で全く新しい発見である。一般論として、申請論文に記載されている成果は、今までほとんど解析のなされていなかった高等生物における構成的な遺伝子発現の機構に対しての研究の基盤を与えるものとして高く評価できる。 申請者は同時にGAPDH遺伝子転写に対するホルモンの作用について調べている。GAPDH遺伝子の転写はインスリンにより上昇することが既に報告されていたが、糖代謝に影響を与えると言われているインスリン以外のホルモンであるグルカゴンとグルココルチコイドの作用について新たに解析し、どちらもGAPDH遺伝子の転写に影響を与えうることを明らかにした。また、未だその作用機構に不明の点の多いインスリンの作用について解析している。まず、インスリンの効果にはグルコースの存在が必要であるかを検討し、GAPDH遺伝子の転写にたいする作用においてはグルコースの存在は必要ではないことを示した。次に、近年多くのインスリン作用がホスホイノシチド3-キナーゼ(以下、PI3Kと略す)により仲介されていることが明らかになっているので、GAPDH遺伝子の転写についてもそうであるかを調べている。申請者はPI3Kに対する特異的な阻害剤であるウオルトマンニンによりインスリンのGAPDH遺伝子発現の上昇効果は打ち消されることを示している。一方でプロテインキナーゼCに対する特異的な阻害剤であるスタウロスポリンはインスリンの作用になんら影響を及ぼさない事も同時に示し、これら事実はからインスリンによる遺伝子転写制御においても、他のインスリン作用同様PI3Kが重要な役割や果たしていると推論している。 以上申請者は本論文においてGAPDH遺伝子のプロモーターの基本構造とホルモン感受性に関して新たな事実を示し、GAPDH遺伝子の発現機構について分子生物学的及び生理学的に重要な知見をつけ加えた。よって本論文は博士(学術)の学位請求論文として合格と認められる。 |