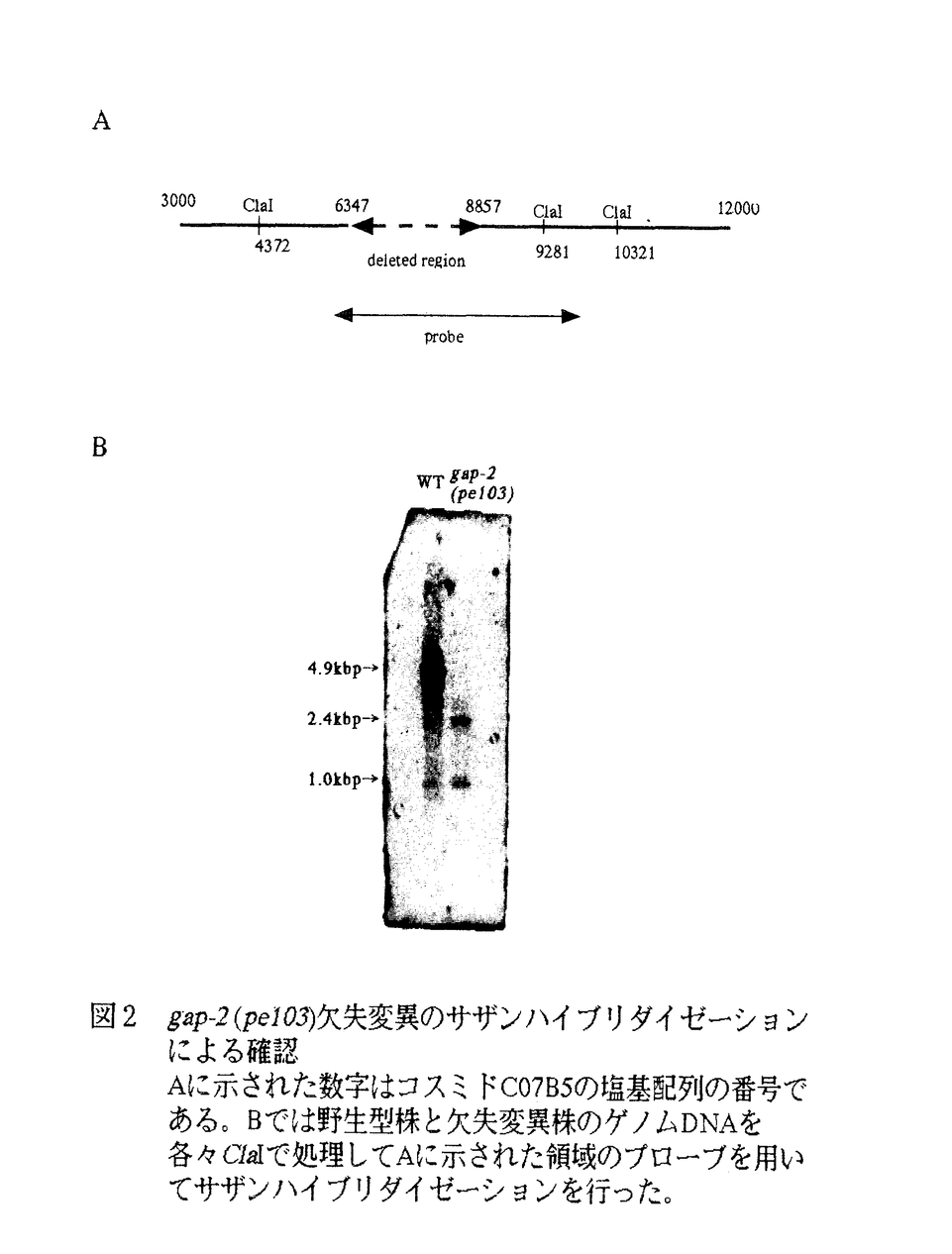
本論文は、線虫Caenorhabditis elegansから新規のRas GTPase活性化因子遺伝子を単離し、性格づけたものである。Rasシグナル伝達経路は広範に保存された経路で、細胞の増殖や分化において中心的な役割を担っている。Rasは細胞膜の内側に存在する低分子量GTP結合タンパク質であり、活性型のGTP結合型と不活性型のGDP結合型をとる。受容体型チロシンキナーゼ等からのシグナルがグアニンヌクレオチド交換因子(GEF)に伝わると、GEFはGDP-RasをGTPRasに変換し、Rasを活性化する。活性化型のGTP結合型Rasは内在性GTP加水分解活性を刺激するGTPase活性化因子(GAP)の作用によってGDP結合型に戻り、シグナルは終結する。C.elegansでは現在までに唯一つのRasであるLET-60が知られている。LET-60を介するシグナルは、発生において陰門形成、排出管細胞形成、オスの交尾針の形成、筋芽細胞の移動、生殖細胞形成過程におけるパキテン期停止解除に必要である。Rasの活性型変異体let-60(n1046)では陰門、排出管細胞の過形成が見られ、Rasの活性調節の重要性を示している。 本論文ではまずRasの活性を負に制御する遺伝子の単離とその機能解析を試みた。分裂酵母内で発現する線虫のcDNAライブラリーを作製し、接合フェロモンに超感受性である分裂酵母のgap1変異を相補するプラスミドpCEGAP3を単離した。挿入されたcDNAから予想されるアミノ酸配列は哺乳類のp120Ras-GAPと最も高い相同性を示したが、先に同定されたC.elegansのRas-GTPase活性化因子遺伝子gap-1とは異なり、それをgap-2と命名した。pCEGAP3を分裂酵母のras1活性化型変異を持つras1Val17変異株に導入しても接合フェロモン超感受性を抑圧できなかったため、gap-2遺伝子は分裂酵母Ras1のGTPase活性を促進することによってRas1の機能を負に制御している可能性が示唆された。 gap-2の全長cDNAを得るために5’-RACE法を行い、複数のバンド中の最長のものとして約900bpのcDNAを取得した。このcDNAは5’端にトランス・スプライシングのリーダー配列であるSL1配列を持ち、全長のcDNAであると判断された。一方、ノザン解析によっても複数種のmRNAの存在が示唆された。SL1リーダー配列を利用したRT-PCRの産物を複数クローニングして塩基配列を決定したところ、さらに8種類のgap-2cDNAが得られた。それらの塩基配列を公開されているC.elegansゲノムの塩基配列と照合したところ、gap-2遺伝子は25個のエクソンからなり、プロモーターの違いやオールタナティブスプライシングにより、少なくとも9つのアイソフォームを生じることが判明した。 gap-2の各アイソフォームのプロモーター領域をそれぞれクローニングし、GFP-lacZレポーターを用いて発現分布を観察した。各アイソフォームは異なる発現分布を示し、アイソフォームが使い分けられている可能性が示唆された。例えば、エクソン4上流域を用いたレポーターは頭部の咽頭筋や咽頭上皮細胞、尾部の芽細胞に強い発現を示し、エクソン13上流域を用いたものは頭部の神経細胞や筋細胞、体側面の下皮細胞や陰門細胞で発現した。in situハイブリダイゼーションでは陰門細胞や生殖腺に強いgap-2mRNAのシグナルが検出された。 Tclトランスポゾンを利用した遺伝子破壊法に従い、C.elegansのgap-2遺伝子破壊株を作製した。ゲノムDNAのサザン解析によりgap-2遺伝子が正しく破壊されていることを確認したが、gap-2単独破壊株において際立った異常は見られなかった。雌雄同体の陰門形成誘導や排出孔細胞の分化誘導に関わるシグナル伝達系では、LET-60Rasの上流に受容体型チロシンキナーゼLET-23が位置しているので、let-23の機能衰退型変異とgap-2遺伝子欠失変異との二重変異を作製して遺伝学的相互作用を調べた。let-23(sy97)変異は幼虫期の致死性(浸透度81.9%)や生き残った成虫の陰門不形成(同100%)という表現型を示すが、二重変異株let-23(sy97);gap-2(pe103)は、成虫の陰門不形成はlet-23(sy97)株と変わりないものの、幼虫期の致死性は減少し、致死性に関してgap-2欠失変異がlet-23(sy97)を抑圧することが示された。let-60(n1876)変異による幼虫期の致死性も同様にgap-2(pe103)変異により部分的に抑圧されることがわかった。いっぽう、RasのエフェクターであるRafの機能衰退型アリルlin-45(sy96)も幼虫期の致死性と成虫の陰門不形成を示すが、lin-45(sy96);gap-2(pe103)二重変異では幼虫期の致死性、成虫の陰門不形成ともに抑圧は見られなかった。これらの観察からGAP-2は幼虫の生育に必須な排出管細胞の誘導においてRasの機能を負に制御していると推定される。gap-2の発現パターン解析によれば、この遺伝子は排出管細胞の誘導以外にも機能を果たしている可能性が高く、本研究で得られたcDNAと突然変異体は今後これらの未知のシグナル伝達系の解析にも有用な材料となることが十分期待できる。以上の成果は発生過程におけるRasシグナル制御系の役割の理解に対して大きな寄与をなすものである。 なお、本論文は飯野雄一および山本正幸との共同研究であるが、論文提出者が主体となって分析および検証を行ったもので、論文提出者の寄与が十分であると判断する。 よって、博士(理学)の学位を授与できると認める。 |