鞭毛運動の原動力となる微小管滑り運動は、微小管上に規則的に並ぶダイニン内腕、外腕が隣接する微小管に対して形成するクロスブリッジの力発生に基づいている。この作用は、ダイニン腕のATP加水分解反応と連関している。ATP加水分解酵素としてのダイニン外腕の性質は詳しく調べられており(Johnson,1985)、その反応機序は、急速なATPとの結合とその加水分解の過程、それに引き続くゆっくりとした分解産物(リン酸およびADP)の放出からなることが知られている。ADPの放出過程は微小管の存在によって加速されることから、ダイニン-ADP複合体が微小管に対する力発生に連関しているのではないかと考えられてきたが、その実験的証拠はなかった。ダイニンの力発生に関わる反応中間体の同定は、鞭毛運動の研究のみならず、モータータンパク質に共通の動作原理を知る上で極めて重要である。本研究の第一部ではウニ精子鞭毛軸糸を材料として、紫外線照射によって加水分解可能なATPを放出するcaged-ATPを用い、急速なATP投与にともなう微小管滑り出し運動の力発生を計測した。この時間経過をこれまでの生化学的な研究から求められたATP加水分解反応の速度と比較することにより、ダイニンの力発生に関わる反応中間体の同定をおこなった。 材料は主としてタコノマクラおよびハスノハカシパンの精子を用いた。精子の細胞膜をTritonX-100を含む溶液で除去した後に、その鞭毛軸糸をホモジナイザーで長さ5-20 m前後の断片にした。これらをカバーガラス上に固定後、10gml-1のエラスターゼを含む溶液で3分間前後の処理をおこなった。実験槽内に2mMのcaged-ATPを含む溶液を導入したのち、弾性係数が0.5-20pN nm-1前後の微小ガラス針を軸糸に付着させた。その後、波長330-400nmの紫外光を試料に10msec照射し、caged-ATPの光分解をおこなった。この実験系におけるATP放出速度の実測値(35.6s-1)及びcaged-ATPの分解効率(約50%)から、2mMのcaged-ATPを用いた場合、試料を中心とした半径150mの領域におけるATP濃度は、紫外線照射開始から数msecで運動開始の閾濃度を越え、10msec前後で、100M、100msecで1mM前後に到達すると考えられる。 m前後の断片にした。これらをカバーガラス上に固定後、10gml-1のエラスターゼを含む溶液で3分間前後の処理をおこなった。実験槽内に2mMのcaged-ATPを含む溶液を導入したのち、弾性係数が0.5-20pN nm-1前後の微小ガラス針を軸糸に付着させた。その後、波長330-400nmの紫外光を試料に10msec照射し、caged-ATPの光分解をおこなった。この実験系におけるATP放出速度の実測値(35.6s-1)及びcaged-ATPの分解効率(約50%)から、2mMのcaged-ATPを用いた場合、試料を中心とした半径150mの領域におけるATP濃度は、紫外線照射開始から数msecで運動開始の閾濃度を越え、10msec前後で、100M、100msecで1mM前後に到達すると考えられる。 紫外線照射によるcaged-ATPの光分解後、鞭毛軸糸は微小管滑り運動を開始するが、この滑り力を軸糸に付着させたガラス針の変位によって計測した。この力計測の結果、鞭毛軸糸は紫外線照射開始から10msecの潜伏時間をおいて力発生を開始し、その値は光照射開始から50-60msecで最大値に到達することが明らかとなった。このようにして得られた力発生の時間変化を、テトラヒメナ22SダイニンのATP加水分解反応における各反応ステップの速度定数(Holzbaur and Johnson,1989)及び本研究の実験系におけるATP放出速度を用いて求めたダイニン-ATP、ダイニン-ADP-Piそしてダイニン-ADP複合体の濃度変化と比較した(図1)。その結果、得られた力発生の時間変化は、微小管によるADP放出速度の加速を考慮に入れたダイニン-ADP複合体生成の時間変化に極めてよく一致する事がわかった。このことはダイニン-ADP複合体がダイニンの力発生を担う反応中間体であることを強く示唆している。  図1 図1 さて、断片化した鞭毛輔糸をカバーガラスに固定し、エラスターゼ処理をおこなわずにATPを投与すると、通常の鞭毛打頻度よりもはるかに高い周波数(〜300Hz)で数nmの振幅を持つ軸糸の振動現象を誘発することが出来る(Kamimura and Kamiya,1989)。この振動現象もダイニンと微小管の相互作用を反映していると考えられているが、その発生機構は不明である。そこで、急速なATP投与に伴う高速微小振動発生の時間経過を調べることにより、この現象とダイニンによるATP加水分解反応との連関について検討した。 その結果、紫外線照射後、鞭毛軸糸の高速振動は微小管滑り出し運動とほぼ同様の潜伏時間を経て再活性化されることがわかった(図2)。しかしながら、微小管滑り出し運動に伴う力発生の時間変化とは異なり、高速振動の周波数は光照射開始から20msec以内にその最大値に到達し、以後ほぼ一定の値となることが明らかとなった。このことは、振動現象にみられる個々の微小管往復運動は、比較的少数のダイニンの運動を反映したものであることを示唆している。  図2 図2 さらに本研究では、ダイニンの力発生とダイニン-ADP複合体生成の連関を詳しく探る目的で、ダイニンの強力な阻害剤であるバナジン酸の効果について検討した。バナジン酸はダイニン-ADPと安定な複合体を作り(Kd=10-20nM)、これらの結合・解離速度は、同様な作用を受けるミオシン-ADP複合体とのそれと比較して106倍速いことが知られている。 そこで、20-80Mバナジン酸存在下で2mM caged-ATPの光分解をおこない、ATPを急速に投与すると、光照射から100-150msecで高速微小振動はほぼ完全に阻害されるが、高速振動の開始時間、振動周波数の最大値および最大値に到達する時間は、バナジン酸の影響を全く受けないことがわかった(図3)。この結果は微小管滑り出し運動に伴う力発生の時間変化についても同様で、力発生の開始時間および最大力発生に到る時間はバナジン酸の存在に影響されなかった。これらの結果は、力発生に関わる反応中間体の形成はバナジン酸による阻害を受けないことを示唆している。 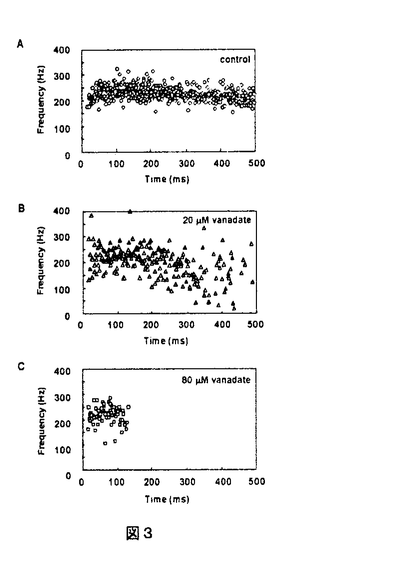 図3 図3 ところが、ダイニン-ADP複合体とバナジン酸の結合・解離の速度定数などから、80Mのバナジン酸存在下では、リン酸が放出された直後、バナジン酸はダイニン-ADPと速やかに複合体を形成すると考えられる。この結果ダイニン-ADP複合体の濃度は著しく低下することが予想される。このことは、前述した「ダイニン-ADP複合体がダイニンの力発生を担う反応中間体である」という結論と矛盾する。 以上の論議をふまえ、本研究では生化学的な研究から指摘されていた、2種類のダイニン-ADP複合体(Holzbaur and Johnson,1989)の存在を仮定した。リン酸が放出された直後のダイニン-ADPはバナジン酸非感受性で、力発生とこれに伴う構造変化を引き起こす。この結果生じたもうひとつのダイニン-ADPはバナジン酸感受性で、これがバナジン酸との速やかな複合体形成をおこなうのではないかと考えている。 さて、鞭毛運動はダイニン腕と微小管の力学的相互作用を原動力とするが、その屈曲波動運動は、鞭毛の長さ方向に並ぶダイニン腕の化学的・力学的活性が高度に制御された結果であると考えられている。しかしながらその制御を担う実体についてはほとんどわかっていない。本研究の第二部では、屈曲波形を保持したままライガー状態にあるウニ精子鞭毛全体に同時に、空間的に一様な濃度のATPを投与し、その波形変化を追跡する事により、ダイニンのATP加水分解サイクルと鞭毛の屈曲波形制御機構の連関を明らかにすることを試みた。 材料は主としてムラサキウニの精子を用いた。除膜した精子のけん濁液を実験槽内に導入し、鞭毛モデルの頭部のみをカバーガラス上に固定した。実験槽内に2mMのcaged-ATPを含む溶液を導入した後、試料に紫外光を照射し、caged-ATPの光分解をおこなった。Caged-ATPの光分解に伴うATP濃度変化は第一部で述べたものと同様である。 再活性化された鞭毛モデルから急速にATPを除去すると、鞭毛モデルは屈曲波形を維持したままライガー状態へと移行する(Gibbons and Gibbons,1974)。このライガー波形鞭毛モデルの屈曲波形が急速なATPの投与に伴って、どのように変化するのかを追跡した。その結果、紫外線照射開始から10-17msecの潜伏時間をおいて、ライガー波形鞭毛は定常的な鞭毛打運動時にみられるものと同様な屈曲伝搬を再開し、逆方向への屈曲伝搬や、波形の弛緩などはみられなかった。このことから、屈曲波形を持つライガー状態の鞭毛内では、ダイニンの滑り活性を制御する仕組みは維持されており、ATPの急速な投与に伴い、ダイニンはそれぞれの制御状態に応じて滑り出し、スムーズな屈曲伝搬が再活性化されると考えられる。 このような再活性化を様々な屈曲形態をとるライガー鞭毛について試みたところ、興味深いことに、鞭毛の中部・後部(鞭毛基部から15-20m以上離れた領域)にあるライガー屈曲波形は、それが定常状態時にみられるものと大きく異なるものであっても、再活性化後、その波形を維持したまま鞭毛後方に伝搬されることがわかった。このことは高粘度溶液中での再活性化時に顕著に認められた(図4)。高粘度溶液中では、鞭毛の屈曲波形は通常の再活性化溶液中でみられるものと大きく異なっており(Brokaw,1966)、その最大滑り角度は通常時にみられるものよりもはるかに大きい。しかしながら、通常の屈曲波形に近い波形を持つライガー鞭毛を高粘度溶液中で急速に再活性化すると、鞭毛の中部・後部に維持されていたライガー波形は、その最大滑り角度を厳密に維持したまま、鞭毛後方に伝搬された。これに対し、鞭毛基部から新たに形成された屈曲波形は高粘度溶液中でみられるものと同様であった。これらのことから、特に鞭毛の中部・後部においては、急速なATPの投与後にみられる微小管の滑りパターンは、その部位に存在したライガー波形に大きく依存することが示唆された。このことは、ダイニンの活性が鞭毛軸糸の形態によって制御されるという仮説を裏付ける一つの傍証と考えられる。  図4紫外線照射(明るい像)から33msecごとの暗視野像を左から右に向かって並べてある。左端はライガー状態の鞭毛像。 図4紫外線照射(明るい像)から33msecごとの暗視野像を左から右に向かって並べてある。左端はライガー状態の鞭毛像。 |