| 内容要旨 | | 進化研究は、近年分子進化学など新しい分野が確立され、様々な知見が得られている。そして、分子進化学において系統樹作成などにより、形態学などの従来の分野で得られてきたデータの、確認および補正・修正がおこなわれ、我々の進化についての認識は、深まりつつある。また、進化理論におけるいくつかの仮説の提唱は、多くの分野の研究を刺激し、大きな影響を及ぼしている。しかし、このような進化についての知見や理論でも解明されない課題が多く残っており、そのなかの一つに、表現型進化の分子レベルからの解明がある。つまり、表現型進化と分子進化の橋渡しである。 分子レベルから表現型進化を考える場合、表現型の形成つまり表現型発現に重要な役割をもつ遺伝子に注目することが、肝要であると思われる。このような表現型発現に重要な遺伝子として、表現型を形成する上で必須の遺伝子調節機構の要となる、転写因子遺伝子がある。そこで私は、この転写因子遺伝子に注目し、その進化を考えることで、分子レベルからの表現型進化の解明をおこなうこととした。そして、その解明の第一段階として、転写因子遺伝子進化とそれによる表現型進化についての仮説の提唱と、その実験的検証を試みた。 遺伝子進化のメカニズムは、現在のところ様々なものが提唱されているが、多くの証拠があり支持を受けているものに、遺伝子重複があげられる。この遺伝子重複は、遺伝子が重複することで新しい遺伝子が生み出せ、機能的分岐を引き起こしうるメカニズムである。しかし、最近、脊椎動物の進化において遺伝子重複は、脊椎動物の進化初期と、有顎脊椎動物の進化初期の二度ほどしか主として生じていないという報告がされている。このため、遺伝子重複以外の遺伝子進化メカニズムが存在すると推測され、この脊椎動物進化における報告から、複雑な有顎脊椎動物の間のような系統的に近い種間で表現型分岐を引き起こしうるメカニズムは、より普遍的に表現型進化を生じさせうるものと考えられる。 ところで、表現型発現に重要な転写因子を様々な種のホモログ間で比較すると、アミノ酸配列の相同性より、保存的な領域と非保存的な領域とが観察される。このことは、転写因子の領域上に、突然変異の蓄積の多かった部分と少なかった部分とが存在し、転写因子の多くが保存領域にDNA結合ドメインをもっていることから、この変異数の違いは、転写因子の祖先において、タンパク内機能間で機能に対する淘汰圧に違いがあったことを意味すると推測される。 そこで、この推測から転写因子の祖先において、淘汰圧の違うタンパク内機能を担う二つの領域があり、その一つは淘汰圧の低い機能を担い、多くの変異の蓄積とそれによる機能的多様化が期待される領域、もう一つは淘汰圧が高い機能を担い変異が起こりにくく、機能的変化が生じにくいと期待される領域だと考えた。そして、前者をLow functional constraint region、後者をHigh functional constraint regionと呼ぶこととし、転写因子ホモログ間での非保存領域は、共通の転写因子祖先のLow functional constraint regionに由来し、保存領域は、High functional constraint regionに由来すると推測した。またさらに、私は、この転写因子祖先が淘汰圧の異なるタンパク内機能を担う二つの領域からなるという概念に基づき、分子レベルからの表現型進化を説明する一つの案として、転写因子の機能的多様化による表現型進化の仮説を提唱した。この仮説は、四つの段階からなり、1.Low functional constraint regionの機能的改変2.転写因子の機能的多様化3.遺伝子カスケードの改変4.生物の形態および生理機能の変化(表現型変化)である。これら概念と仮説は、全ての遺伝子の進化および表現型進化に適用できるわけではないと思われるが、近縁な種間での表現型の違いの由来を説明しうる可能性があるものであり、分子進化と表現型進化を橋渡しする普遍的な一つのメカニズムを提示しうるものと考えている。 科学的仮説は必ず検証することができなければならず、またその必要がある。このため私は、自分の提唱した概念と仮説を検討するための実験をおこなった。 提唱した概念と仮説の実験的な検証には、方法的にいくつかあると思われるが、私は、二つの段階からなる方法をとった。それは、まず第一に、進化の過程で段階的に変化している表現型に重要な転写因子ホモログを、共通の基本設計をもつ種同士から単離の後、その配列を比較し、共通転写因子祖先のLow functional constraint regionに由来すると思われる領域の同定をおこない、第二に、同定された領域の機能を種間で比較するというものである。私は遺伝子重複が、有顎脊椎動進化の大半では主として生じていないことから、表現型を有顎脊椎動物進化で段階的に変化している脳機能とし、その脳機能が爬虫類と哺乳類の間で、大脳大型化を伴って大きく改変されているため、比較動物として、爬虫類と哺乳類がよいと考えた。そして、爬虫類として、爬虫類綱の中で種の多いトカゲ亜目の一種であるアメリカカメレオン(学名Anolis carolinensis英名green anole)を、哺乳類として、脳機能の目覚ましい変化をとげたヒト(学名Homo sapiens)を用いることとした。また、対象とする転写因子は、主として脳神経系に発現をし、脳機能の分化・発現に非常に重要な役割をもつclass III POUとした。 アメリカカメレオンclass III POU遺伝子の単離は、アメリカカメレオンゲノムDNAライブラリーのスクリーニングにておこない、今回class III POU遺伝子の一つであるBrain-1遺伝子の部分的クローンを得ることができた。そして、このクローンは、すでに報告されているアメリカカメレオンBrain-1の完全な配列と全く同一のものであり、系統樹作成による解析によりorthologousなものであると考えられた。このアメリカカメレオンBrain-1と、すでに得られているヒトBrain-1との間でアミノ酸配列の比較をおこなったところ、N末領域に違いがみられた。また、Brain-1遺伝子は、マウス・ゼブラフィッシュからも単離されており、ヒトとアメリカカメレオンを加え四種の動物間でアミノ酸配列を比較したところ、やはりN末領域にアミノ酸配列の違いがみられ、N末領域は非保存的な領域であると考えられるとともに、それぞれのN末領域は、Brain-1の祖先の同じLow functional constraint regionに由来していると推測された。 そこで次に、ヒトとアメリカカメレオンの間で、Brain-1ホモログのN末領域の機能の比較をおこなった。Brain-1は転写因子であり、DNA結合ドメインであるPOUドメインがN末領域内にないことから、N末領域の機能として、転写活性化能が推測された。このため、転写活性化能を測定することができる、哺乳類細胞でのGAL4を利用したルシフェラーゼアッセイを、機能比較のため用いることとした。また、このルシフェラーゼアッセイは哺乳類細胞を使用したものであるので、ヒトBrain-1のN末領域に対する哺乳類特異的コファクターの関与が予測された。そこで、この点に関する情報を得る目的で、ヒトともアメリカカメレオンとも系統的に遠い真核生物である酵母でのGAL4を用いた -ガラクトシダーゼアッセイもおこなうこととした。 -ガラクトシダーゼアッセイもおこなうこととした。 この結果、哺乳類でのアッセイにおいて、アメリカカメレオンBrain-1のN末領域の方がヒトBrain-1のN末領域よりも高い活性化能を示し(図1)、この傾向は、酵母でのアッセイでも同様に検出された(図2)。 これら種間での活性化能の相違は、共通なBrain-1祖先内のLow functional constraint regionに、進化の過程で機能的変化が生じてきたことを強く示唆する証拠の一つとなると考えられ、私の提唱した概念と仮説を支持する一つのよりどころとなると思われる。また、私の概念と仮説を支持するとみられるいくつかの報告があり、提唱した概念と仮説は、広く表現型進化を説明できるものではないかと考えられる。 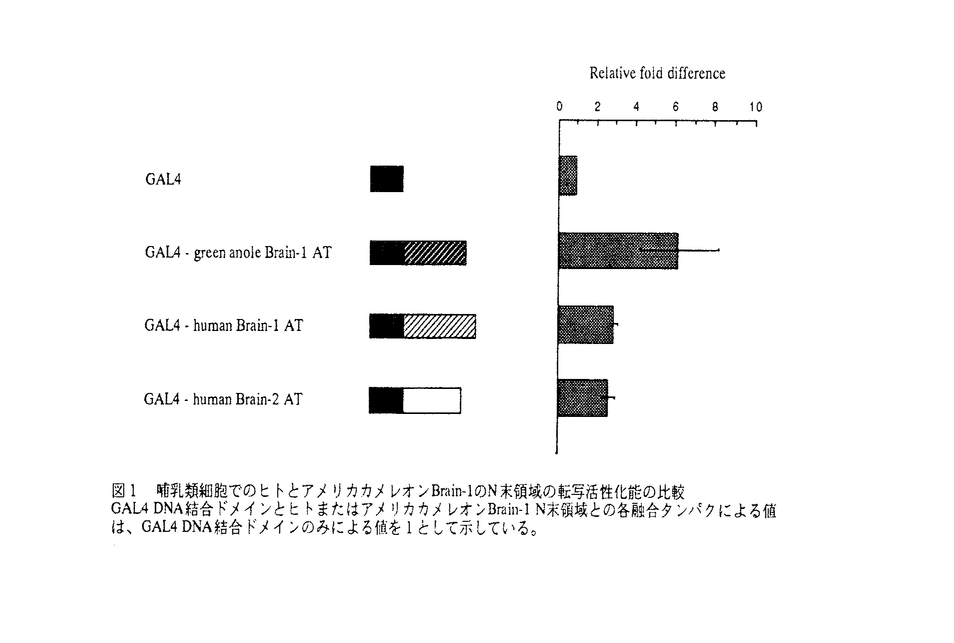 図1 哺乳類細胞でのヒトとアメリカカメレオンBrain-1のN末領域の転写活性化能の比較GAL4 DNA結合ドメインとヒトまたはアメリカカメレオンBrain-1N末領域との各融合タンパクによる値は、GAL4 DNA結合ドメインのみによる値を1として示している。 図1 哺乳類細胞でのヒトとアメリカカメレオンBrain-1のN末領域の転写活性化能の比較GAL4 DNA結合ドメインとヒトまたはアメリカカメレオンBrain-1N末領域との各融合タンパクによる値は、GAL4 DNA結合ドメインのみによる値を1として示している。 図2 酵母でのヒトとアメリカカメレオンBrain-1のN末領域の転写活性化能の比較-ガラクトシダーゼ酵素活性は、酵素量にて示している。 図2 酵母でのヒトとアメリカカメレオンBrain-1のN末領域の転写活性化能の比較-ガラクトシダーゼ酵素活性は、酵素量にて示している。 |
| 審査要旨 | | 本論文は,中枢神経系において遺伝子の発現を特異的に制御している転写因子class III POUのメンバーの一つであるBrain-1の転写活性化領域であるN末領域に関して,ヒト(哺乳類)とアメリカカメレオン(爬虫類)の間での一次構造(アミノ酸配列)上および機能(転写活性可能)上の相違,について述べられている。 まず,アメリカカメレオン(Anolis carolinensis)ゲノムDNAライブラリーよりBrain-1遺伝子の単離がおこなわれた。遺伝子領域を含むDNA断片をプラスミッドベクターにサブクローニングした後,両鎖の配列決定がなされ,その遺伝子構造が明らかにされている。アメリカカメレオンBrain-1遺伝子とorthologousな関係にあるヒトのBrain-1遺伝子との間でアミノ酸配列の比較がおこなわれ,両者の間にはN末端領域に著しい違いが存在することが明らかにされた。Brain-1遺伝子は,マウス・ゼブラフィッシュからも単離されており,ヒトとアメリカカメレオンを加えた4種類の脊椎動物間でアミノ酸配列を比較したところ,同様にN末端領域で著しいアミノ酸配列の違いがみられ,N末端領域は進化的に非保存的な領域であることを明らかにした。 次に,ヒトとアメリカカメレオンの間で,Brain-1ホモログのN末端領域の機能の比較がおこなわれている。転写因子Brain-1のDNA結合ドメインはPOUドメインと呼ばれるC末端側の領域にあり,N末端側の領域は転写活性化能を担っていると考えられている。すなわち,遺伝子発現量(どれほどの遺伝子量の発現を引き起こすができるか)に関与する領域と考えられている。そこで,転写活性化能を比較測定することができる実験系としてルシフェラーゼ遺伝子をレポーター遺伝子としたone hybrid systemを用いることによって解析をおこなった。すなわち,酵母の転写因子であるGAL4のDNA結合ドメインにヒトならびにアメリカカメレオンそれぞれのBrain-1遺伝子のN末端領域をつなげ,この融合蛋白を発現させるために発現ベクターに組み込む。これらをそれぞれ独立に,レポーター遺伝子であるルシフェラーゼ遺伝子の上流に酵母の転写因子GAL4の結合配列をもつレポーターベクターと共に哺乳類培養細胞にエレクトロポレーション法により導入している。このようにして構築した系をもちいて,ヒトとアメリカカメレオンの間での,Brain-1ホモログのN末端領域の転写活性化能の比較解析がおこなわれ,両者の転写活性化能には少なくとも2倍相当の違いが存在することを明らかにしている。なお,本論文は哺乳類と爬虫類の間での違いを解析することを目的に実験分析をすすめているが,用いた上記の系は哺乳動物由来の培養細胞を含む要素から構成されている。そこで,哺乳動物に特異的な転写仲介因子の存在を原因として得られた結果である可能性を検討(排除)することを目的として,ヒトならびにアメリカカメレオンの両者から系統的に遠い(同距離にある)酵母を哺乳動物の培養細胞に替わる系としてもちいた同様の実験をおこなっている。得られた結果は上記と全く同様の結果であり,ヒトとアメリカカメレオンの間でBrain-1遺伝子N末端領域の転写活性化能に違いが存在すること(すなわち,用いた実験系を問わず同様の違いが存在すること)を確認している。 以上の結果は,ヒト(哺乳類)とアメリカカメレオン(爬虫類)の間でみられた転写因子Brain-1の活性化能の相違は,進化的に保存性の低いN末端領域の違いによるものであることを示している。従来転写因子の強い進化的保存性が言われてきたが,転写因子の非保存的領域の存在とそれが機能的差異を生じさせていること,が本論文によって初めて明らかとなった。 なお,本論文は大石直哉・植田信太郎との共同研究であるが,論文提出者が主体となって分析及び検証を行ったもので,論文提出者の寄与が十分であると判断する。 よって,博士(理学)の学位を授与できると認める。 |