| 内容要旨 | | 「序論」 マメ科植物-根粒菌共生相互作用は、高等植物と細菌間で成立する細胞内共生というユニークかつ複雑な生物間相互作用である。しかしながら、窒素固定の場となる根粒の形成過程に直接関与する植物側の共生関連遺伝子及びその分子機構についての知見は殆ど得られていない。私は、研究対象として、マメ科植物であるミヤコグサLotus japonicusと、その共生菌であるMesorhizobium lotiとの共生系を選んだ。ミヤコグサは、ゲノムサイズが小さく、1世代期間が約3ヶ月、Agrobacteriumによる形質転換及び個体の再分化が容易である(表1)など、分子遺伝学的解析に好適な形質を有し、国内外でもマメ科のモデル植物候補として活発な研究の対象となりつつある。このミヤコグサを用い、栽培・交配・in vitro根粒菌感染系などの基本実験系の検討・確立を行うと共に、将来的に植物側の根粒器官形成の分子機構を解明することを目的として、根粒菌との初期認識過程から窒素固定活性発現に至る過程に異常を示す変異体を11系統単離することに成功した。このうち、特に窒素固定活性発現過程に異常を示す二つの変異体、alb1,fen1に的を絞り、これら変異体の共生成立破綻が、植物側の根粒菌に対する異常な防御応答発現、あるいは根粒器官形成過程の異常に起因している可能性についての検討・解析を行った。 「実験方法と結果」1.EMS処理による根粒形成過程に異常を示すミヤコグサ変異体の単離及び遺伝解析 ミヤコグサ(Lotus japonicus,B-129;Gifu)種子についてEMS処理を行い、得られた約50,000個のM2種子のうち17,000個を用い、共生変異体のスクリーニングを行なった結果、11系統の共生変異体の単離に成功した。これらのうち、現時点において、monogenicかつrecessiveな遺伝性を示す変異体として、根粒非着生(Nod)の表現型を示すnss1,lul1変異体(Nodカテゴリーにおいて、相互にnon-allelic)、根粒器官形成から窒素固定活性発現過程に異常を示すalb1,fen1変異体(Nod+Fix-;同カテゴリーにおいて、相互にnon-allelic(表2))の4遺伝子座を同定している。 2.根粒器官形成から窒素固定活性発現過程に異常を示す(Nod+Fix-)変異体--alb1,fen1の表現型解析--(1)alb1(aberrant localization of bacteria inside nodule)変異体 alb1変異体は、根粒形成の遅延が認められると共に、いびつな形状の大小2種類の根粒を着生する。大多数の根粒(first type)は白色・小型であり、窒素固定活性を有さない(図1左)。長期感染実験の継続により低頻度ながら着生する大型根粒(second type)は、窒素固定活性を有する。first type根粒では、根粒菌を含む感染細胞が認められない。さらに、細胞間隙を押し広げるような構造の内部に多数の根粒菌が局在する様子が観察された事から(図3B,3D)、この変異体をalb1(aberrant localization of bacteria inside nodule)と命名した。さらにalb1変異体では、根粒菌の侵入経路となる感染糸内が異常肥大し、その内部に多数の根粒菌が圧縮された状態で存在していた。一方、second type根粒の内部には、窒素固定活性を示す根粒菌に特異的な形態であるバクテロイドに分化した根粒菌を含む巨大な感染細胞(図3C,E)が認められた。従って、first typeの根粒のうち、低頻度ながら宿主細胞への侵入に成功したものがsecond typeの根粒に発達したと考えられる。また、alb1根粒では、根粒維管束鞘の分化・発達不全の異常が認められた(図4A,B)。これらの結果より、alb1変異体は、根粒菌の宿主細胞への侵入過程及び根粒器官形成に異常を示す変異体であると考えられる。 (2)fen1(fail in enlargement of infected cells of nodule)変異体 fen1変異体は、wild type(以下wt)根粒と比較して、小型で薄桃色の根粒を着生する(図1右)。これらの根粒は着生後数日を経て薄緑色に変色し、微弱な窒素固定活性を有するものの、その活性が速やかに低下することからpre-mature senescenceタイプの変異体であると考えられる。根粒内部形態の観察より、fen1ではバクテロイドに分化した根粒菌を含む多数の感染細胞が認められたが、これらの細胞は肥大成長できず(図2B)、速やかに細胞質の崩壊が進行する(図2C)ことから、この変異体をfen1(fail in enlargement of infected cells of nodule)と命名した。 3.alb1,fen1変異体における宿主防御応答関連遺伝子の発現解析 根粒菌の細胞内共生及び根粒の窒素固定活性発現に異常をきたす変異体の中には、感染・侵入してくる根粒菌を「異物」と見なして、植物病原菌(非親和性菌)に対して発現する防御応答に類似した反応を引き起こしている例が報告されてる。alb1,fen1変異体において、このような異常な防御応答発現が共生成立破綻の原因となっている可能性を検討するため、ミヤコグサcDNA libraryよりクローニングされた、phytoalexin合成を司るphenylpropanoid pathwayのkey enzymeであるchalcone synthase(CHS)、chalcone isomerase(CHI)及びisoflavone reductase(IFR)遺伝子を指標としたNorthern hybridizationを行った。しかしながら、現時点で、防御応答発現を強く示唆するデータは得られていない。 4.alb1,fen1変異体におけるnodulin遺伝子の発現解析 変異体における根粒形成過程の異常を、根粒器官形成特異的に発現するnodulin遺伝子の発現レベルから解析することを目的として、根粒形成初期から根粒器官形成過程を通じて発現する初期nodulinであるLjenod40遺伝子、及び根粒形成後期の窒素固定活性発現過程に発現する後期nodulinであるLjLb遺伝子を指標としたNorthern及びin situ hybridizationによる解析を行った。fen1変異体では、Ljenod40及びLjLbの発現は微弱ながら認められたが、その発現量は速やかに低下しており(図5)、nodulin遺伝子発現レベルにおいてもpre-mature senescenceタイプの変異体であることが強く示唆された。一方alb1変異体ではLjenod40及びLjLbの発現はNorthern hybridizationでは認められなかった(図6)。そこで根粒内部でのLjenod40mRNAの局在・発現解析を目的として、wt及びalb1根粒についてin situ hybridizationによる観察を行った。wtでは、根粒形成初期には根粒原基近傍のpericycleに(図7A)、成熟した根粒では根粒維管束鞘を取り囲むparenchyma領域及び非感染細胞にその発現が強く認められた(図7C)。一方alb1変異体では、極めて微弱なLjenod40の発現がPericycle(図7B)及びparenchyma(図7D)で認められた。従って、alb1変異体の根粒器官形成過程において、Ljenod40遺伝子の発現が著しく抑制されていると考えられる。 「考察及び展望」 本研究は、根粒形成初期から窒素固定活性発現に至る根粒器官形成プロセスの宿主制御因子を分子レベルで解明することを目的として行ったものであり、ミヤコグサLotus japonicusの共生変異体を用いた分子遺伝学的解析に向けての最初に位置する研究である。私は、本研究において単離したalb1,fen1変異体について、共生成立の破綻原因を検討するため、(1)根粒菌に対する宿主の異常な防御応答発現、(2)根粒器官形成進行過程の異常、の二方向からの解析を試みた。(1)においては、防御応答遺伝子の指標としたCHS及びCHI,IFR遺伝子の発現解析より、防御応答の発現を強く示唆する結果は得られていない。しかしながら、(2)においては、極めて興味深い結果が得られた。Nodulin遺伝子の発現解析より、alb1変異体では根粒器官形成時のLjenod40遺伝子の発現が著しく抑制されていることが明らかとなった。enod40遺伝子は、これまで調べられた全てのマメ科植物で良く保存されており、根粒原基を構成する皮層細胞の細胞分裂に先立ってpericycleで誘導され、根粒器官形成特異的に発現する。また、根粒菌の感染を伴わずに形成されるspontaneous noduleや根粒菌の共生変異株により誘導されるempty noduleなど、共生成立を伴わずに根粒器官が形成される無効根粒においても発現することが報告されている。さらに、その機能解析より、enod40遺伝子は、根粒菌との初期認識過程で早期に発現し、細胞中のホルモンバランスを変化させることにより、根粒原基となる皮層細胞の細胞分裂を誘導していると推定されている。従って、alb1変異体における根粒形成の遅延・根粒維管束鞘の発達不全及び感染糸発達異常とenod40遺伝子の発現抑制の関連性に興味が持たれる。さらに、皮層細胞分裂誘導に加え、根粒維管束鞘の発達・分化においてもenod40遺伝子が重要な役割を担っている可能性も考えられ、根粒器官形成におけるenod40の役割をさらに解析する上で、alb1変異体は有効な系となりうるものと考えられる。 EMS処理により得られた残りの33,000個のM2種子についてもスクリーニングを行った結果、さらに複数の共生変異体の単離に成功し、それらの遺伝・表現型解析が進行中である。これらの変異体の表現型をもとに根粒器官形成過程を細分化し、細分化されたそれぞれの過程に関与する宿主の共生関連遺伝子を解明する基盤づくりが進みつつある。  表.1 ミヤコグサの分子遺伝学的解析に適した形質 表.1 ミヤコグサの分子遺伝学的解析に適した形質 表.2 Gifu(wt)との戻し交配によるalb1,fen1変異体の遺伝解析及びアレリズムテスト 表.2 Gifu(wt)との戻し交配によるalb1,fen1変異体の遺伝解析及びアレリズムテスト 図.1 ミヤコグサ菌Mesorhizobium loti JRL501接種40日後の根粒着生根。写真左側よりalb1,Gifu(wt),fen1。 図.1 ミヤコグサ菌Mesorhizobium loti JRL501接種40日後の根粒着生根。写真左側よりalb1,Gifu(wt),fen1。 図.2 Gifu(wt)及びfen1変異体根粒の感染細胞領域の光学顕微鏡写真。A.Gifu根粒感染細胞領域。肥大成長した感染細胞が存在する。B.fen1の感染細胞領域。感染細胞はGifu(wt)に比べて小型で、老化進行の指標となる感染細胞内の小型液胞(矢尻)及び感染細胞核の濃縮化(矢印)が観察される。C.fen1根粒。急速な老化進行に伴う感染細胞崩壊(矢印)が観察される。(bar=25 図.2 Gifu(wt)及びfen1変異体根粒の感染細胞領域の光学顕微鏡写真。A.Gifu根粒感染細胞領域。肥大成長した感染細胞が存在する。B.fen1の感染細胞領域。感染細胞はGifu(wt)に比べて小型で、老化進行の指標となる感染細胞内の小型液胞(矢尻)及び感染細胞核の濃縮化(矢印)が観察される。C.fen1根粒。急速な老化進行に伴う感染細胞崩壊(矢印)が観察される。(bar=25 m(A,B),50m(C)) m(A,B),50m(C)) 図3 Gifu(wt)及びalb1変異体の根粒内部形態の光学及び電子顕微鏡による観察(菌接種後60日)A Gifu根粒、多数の感染細胞が認められる.B alb1のfirst type根粒、感染細胞は認められず、細胞間隙を押し広げるような構造(矢印)が観察される,C alb1のsecond type根粒、巨大感染細胞が根粒内部に散在している。D Bの細胞間隙のTEMによる観察像、不定形状を示す物質(矢印)と共に多数の根粒菌(矢尻)が局在している.E Cの感染細胞内部。ペリバクテロイド膜(矢尻)にくるまれ、バクテロイドに分化した根粒菌が多数観察される.(bar=50m(A.C).25m(B),2m(D).1m(E)) 図3 Gifu(wt)及びalb1変異体の根粒内部形態の光学及び電子顕微鏡による観察(菌接種後60日)A Gifu根粒、多数の感染細胞が認められる.B alb1のfirst type根粒、感染細胞は認められず、細胞間隙を押し広げるような構造(矢印)が観察される,C alb1のsecond type根粒、巨大感染細胞が根粒内部に散在している。D Bの細胞間隙のTEMによる観察像、不定形状を示す物質(矢印)と共に多数の根粒菌(矢尻)が局在している.E Cの感染細胞内部。ペリバクテロイド膜(矢尻)にくるまれ、バクテロイドに分化した根粒菌が多数観察される.(bar=50m(A.C).25m(B),2m(D).1m(E))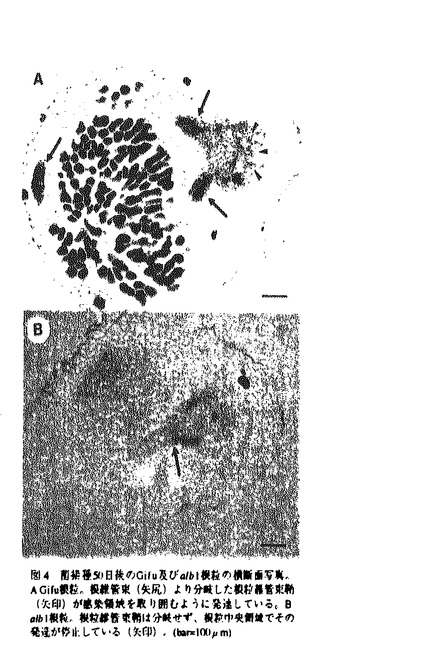 図4 菌接種50日後のGifu及びalb1根粒の横断面写真。A Gifu根粒。根維管束(矢尻)より分岐した根粒維管束鞘(矢印)が感染領域を取り囲むように発達している。B alb1根粒。根粒維管束鞘は分岐せず、根粒中央領域でその発達が停止している(矢印),(bar=100m) 図4 菌接種50日後のGifu及びalb1根粒の横断面写真。A Gifu根粒。根維管束(矢尻)より分岐した根粒維管束鞘(矢印)が感染領域を取り囲むように発達している。B alb1根粒。根粒維管束鞘は分岐せず、根粒中央領域でその発達が停止している(矢印),(bar=100m) 図5 Gifu及びfen1根粒におけるLjenod40及びLiLb遺伝子の発現解析(Northern hybridization)。Leaf、uninfected root及びGifu根粒(接種後10,14,20,30日後),fen1根粒(接種後14,30日後)より抽出したtotal RNA5gをサンプルとした。 図5 Gifu及びfen1根粒におけるLjenod40及びLiLb遺伝子の発現解析(Northern hybridization)。Leaf、uninfected root及びGifu根粒(接種後10,14,20,30日後),fen1根粒(接種後14,30日後)より抽出したtotal RNA5gをサンプルとした。 図6 Gifu及びalb1根粒におけるLjenod40及びLjLb遺伝子の発現解析(Northern hybridization).Leaf、uninfected root及びGifu根粒(接種後10,14,20,30日後),alb1根粒(接種後14,20,30日後)より抽出したtotal RNA 5gをサンプルとした。alb1根粒では、Ljenod40及びLjLb共に発現が認められない。 図6 Gifu及びalb1根粒におけるLjenod40及びLjLb遺伝子の発現解析(Northern hybridization).Leaf、uninfected root及びGifu根粒(接種後10,14,20,30日後),alb1根粒(接種後14,20,30日後)より抽出したtotal RNA 5gをサンプルとした。alb1根粒では、Ljenod40及びLjLb共に発現が認められない。 図7 in situ hybridizationによるGifu(wt.A.C.)及びalb1(B.D)根粒におけるLjenod40 mRNAの根粒器官内の発現パターン解析。(A,B)根粒形成初期の発現パターン。Gifu、alb1共に、根粒原基基部に位置するpcricycle(矢印)に発現が認められるが、alb1(B)のシグナルはGifu(A)に比べ微弱である。(C,D)成熟した根粒内の発現パターン.Gifu(C)では、根粒維管束鞘(矢印)、parenchyma(矢尻)及び非感染細胞領域に強い発現が認められる.alb1(D)では、極めて微弱な(写真では確認できない)シグナルが発達不全の根粒維管束鞘(矢印)周辺にのみ認められた。(bar=50m(A,B),25m(C,D)) 図7 in situ hybridizationによるGifu(wt.A.C.)及びalb1(B.D)根粒におけるLjenod40 mRNAの根粒器官内の発現パターン解析。(A,B)根粒形成初期の発現パターン。Gifu、alb1共に、根粒原基基部に位置するpcricycle(矢印)に発現が認められるが、alb1(B)のシグナルはGifu(A)に比べ微弱である。(C,D)成熟した根粒内の発現パターン.Gifu(C)では、根粒維管束鞘(矢印)、parenchyma(矢尻)及び非感染細胞領域に強い発現が認められる.alb1(D)では、極めて微弱な(写真では確認できない)シグナルが発達不全の根粒維管束鞘(矢印)周辺にのみ認められた。(bar=50m(A,B),25m(C,D)) |
| 審査要旨 | | 共生窒素固定の場であるマメ科植物の根粒は、根粒菌とその根粒菌の宿主となるマメ科植物との緊密な相互作用によって形成される。その相互作用の分子機構の解析は、近年根粒菌側で急速な進展をみせたが、植物側の解析はほとんど進んでいないのが現状である。その一つの原因は、有効な解析手段である分子遺伝学的解析に適したマメ科植物が知られていなかったことにある。本論文は、最近マメ科植物の分子遺伝学的解析に好適な形質を有し、モデル植物の候補として国内外で活発な研究対象になりつつあるミヤコグサ(Lotus japonicus)を用い、根粒形成過程の解析を試みたもので、5章からなる。 第一章ではミヤコグサを用いた遺伝解析を進めるにあたって必要な基本的諸条件を検討し、安定した根粒着生に適した栽培条件の設定、効率のよい交配方法の確立、in vitroでの根粒形成の検定系の改良、ごく初期の根粒形成過程解析のためのspot inoculation法の改良などについて述べられている。 第二章ではEMS処理により得られた根粒形成に異常を示すミヤコグサ変異体の単離とその遺伝解析について記載されている。17,000個のM2種子を用い共生変異体のスクリーニングを行い、遺伝的に安定な11系統の変異体を得た。このうち遺伝解析が進み、monogenicかつrecessiveな遺伝性を示す根粒非着生変異体2系統、窒素固定活性発現過程の変異体(Fix-)2系統を得ている。 第三章では 上記Fix-2系統の表現形の解析を、主として根粒の形状、窒素固定活性、光学顕微鏡および電子顕微鏡による観察によって行なった結果が述べられている。第一の変異体では、根粒形成が遅延し、いびつな形状の大小2種の根粒を着生する。大多数の根粒は白色・小型で窒素固定活性を示さない。また、感染細胞が認められず、細胞間隙や異常肥大した感染糸に、根粒菌が圧縮された状態で多数局在している様子が観察された。大きい方の根粒は少数であるが正常な感染細胞を有しており、稀に根粒菌が侵入に成功した細胞は正常に感染細胞へ分化できると推定された。これらの結果より、この変異体は根粒菌の宿主細胞への侵入過程に異常を示す変異体と推定され、根粒菌が異常な局在を示すことからalb1(aberrantlocalization of bacteria inside nodule)と命名された。 もう一つの変異体は小型・薄桃色の根粒を着生するが、その根粒はすぐ薄い緑色に変色し、僅かに認められていた窒素固定活性も急速に低下することから、いわゆるpre-mature senescence typeの変異体と考えられる。バクテロイドを含む正常に分化した感染細胞は肥大成長できず、速やかな細胞質の分解がおきる。この観察からこの変異体はfen1(fail in enlargement of infected cells of nodule)と命名された。 第四章には、防御応答の変異である可能性について検討された結果を記載されている。根粒形成変異体の中には根粒菌を異物とみなして、非親和性の植物病原菌のように防御応答に類似した反応を引き起こすものがあることが期待される。そこでフィトアレキシン合成を司るchalcone synthase,chalcone isomerase,isoflavone reductaseをプローブとして転写活性を検討したが、異常な防御応答反応を示唆する結果は得られていない。 第五章では上記変異体で見られる根粒形成過程の異常を,根粒特異的に発現するノジュリン遺伝子の発現レベルから解析することを目的として行なわれた結果について述べられている。初期ノジュリンであるLjenod40遺伝子と後期ノジュリンであるレグヘモグロビン(LjLb)遺伝子を指標としてNorthernおよびin site hybridizationによる解析を行なった。fen1変異体ではLjenod40およびLjLbの微弱な発現が認められたが、その発現量は急速に低下し、ノジュリン遺伝子の発現レベルにおいてもpre-mature senescenceタイプの変異体であることが示唆された。Ljenod40は野生型の植物ではまず根粒原基近傍のpericycleで発現し、根粒の維管束の発達につれ、維管束を取り囲む柔組織で発現が継続して見られるが、alb1変異体ではNorthern hybridizationでは発現は認められず、in site hybridizationでpericycleと発達不全の根粒維管束を取り囲む柔組織で微弱ながら発現が認められた。alb1変異体は根粒器官発達過程を通してLjenod40遺伝子の発現が著しく抑制されている新規の変異体であることが示された。 これらの結果の一部は世界で初めてのミヤコグサの変異体として報告されており、根粒形成分子機構の解析に寄与するところは大きい。本論文は全体を通して共同研究で行なわれているが、論文提出者が主体になって計画の立案し、実験を行なったものである。 したがって、博士(理学)の学位を授与できると認める。 |