| 内容要旨 | | 光合成のCO2固定酵素であるribulose-1,5-bisphosphate carboxylase/oxygenase(Rubisco)は、通常の陸上植物では葉緑体ストロマ中に分散しているが、ツノゴケの一部と大部分の真核藻類の葉緑体内では集合して基質構造をとり、スターチに囲まれ『ピレノイド』として存在している。しかしながら、ピレノイド構造の生物学的意義については不明な点が多く、その生理的役割を直接的に証明した研究例はない。Rubiscoは最大反応速度が遅く、CO2に対する親和性が低いなどの特性を持つ酵素であるが、藻類では、細胞としてのCO2に対する親和性は高くなっており、この現象はCO2をRubiscoの回りに集める『CO2濃縮機構』が働いていることで説明されている。CO2濃縮機構には、carbonic anhydrase(CA)、細胞内への無機炭素の蓄積に加え、ピレノイドも関与することが示唆されてきている。 一方、藻類の中にはピレノイドを持たない種もあり、ピレノイドを持つ種においても、その形態は多様である。近年、地衣類の共生藻を用いた比較生理学的研究が行われ、ピレノイド構造の生理的役割が議論された(e.g.Palmqvist et al.1994)。これらの研究では、用いたピレノイドを欠く藻類数種すべてがCO2濃縮機構を欠いており、ピレノイドがCO2濃縮機構に必須であると議論された。しかしここで調査された藻類は地衣類の共生藻に限られており、近縁なピレノイドを持つ種との比較もなされていない。 本研究では、真核藻類で最もCO2濃縮機構の研究の進んだ種の一つであるChlamydomonas(Cd.)reinhardtiiが所属するChlamydomonas属に近縁でかつピレノイドを欠くとされる自由生活藻Chloromonas(Cr.)属、並びにChlamydomonas属を材料とし、その葉緑体のピレノイドに関する微細形態とCO2濃縮機構を比較し、ピレノイドの機能を推測することを目的とした。この中で、用いた株の系統解析を行うことにより、解析結果に基づく近縁な生物間での比較生物学的研究が可能となった。 <結果および考察>1.ピレノイドにおけるRubiscoの集中 Suss et al.(1995)は凍結固定法とホウレンソウのRubisco抗体を用いた免疫電子顕微鏡(電顕)法による研究結果から、Cd.reinhardtiiのRubiscoの大部分はピレノイドではなくストロマに存在していると報告した。これはピレノイドがRubiscoの集中した構造である、という本研究の前提と矛盾するため、Cd.reinhardtiiのRubisco large subunit抗体を用いて再検査した。その結果、凍結法を用いても従来の常温での固定法と同様の結果が得られ、Cd.reinhardtii細胞でRubiscoがピレノイドに集中していることが再確認された。 2.ピレノイドを欠くChloromonas株の形態及びCO2濃縮機構 カルチャーコレクションで保存されているChloromonas株の中から、光学顕微鏡と透過型電顕でピレノイド構造のないことが確認され(図1)、さらに免疫電顕で、葉緑体内にRubiscoの集中した部位が認められなかった、Cr.serbinowii U492、Cr.rosae U1337、Cr.clatharata U1970、Cr.rosae S26.90、Cr.palmelloides S32.86でCO2濃縮機構の検討を行った。多くの微細藻類において、1-5%のCO2を通気して培養した細胞(高CO2細胞)に比べ、通常の空気を通気して培養した細胞(低CO2細胞)で、CO2に対する親和性(細胞の光合成酸素最大発生量の1/2を与えるCO2濃度、K0.5(CO2)で示す)が高くなるが、Rubiscoの親和性はCO2条件により大きく変化しないため、この現象は、CA活性の誘導などCO2濃縮機構の発現によるものと考えられる。そこで、異なるCO2条件で生育させた細胞でCO2に対する親和性とCA活性を調査し、さらに細胞内に蓄積された無機炭素の濃度を調査した。その結果、上記5株は2つのタイプに分かれた。Cr.serbinowii U492とCr.rosae U1337では、低CO2細胞で高CO2細胞に比べ、細胞のCO2に対する親和性が高く、低CO2細胞でCA活性が誘導されており、細胞内に無機炭素が蓄積されていた(表1)。従って、これらの株ではピレノイドがないにもかかわらず、CO2濃縮機構を保持していることが明らかになった。一方、Cr.clatharata U1970、Cr.rosae S26.90、Cr.palmelloides S32.86では、CO2に対する親和性は、高CO2細胞でも低CO2細胞でも低く、CA活性は高CO2細胞と低CO2細胞の間で差が認められず、細胞内に無機炭素が蓄積されていなかった(表1)。従ってこれらは地衣類のピレノイドのない共生藻と同様、CO2濃縮機構を持たないことが判明した。ピレノイドがないにもかかわらずCO2濃縮機構を保持する藻類の存在は、これが初めての報告である(表3)。ただし、これら2株ではCd.reinhardtiiに比べると無機炭素の蓄積は1/10程度であり(表1)、無機炭素の蓄積の大きさに、ピレノイドの存在が関係している可能性が考えられた。 3.Chloromonasに近縁なChlamydomonas等の株の形態及びCO2濃縮機構 上述のピレノイドのないChloromonas属に近縁とされるChlamydomonas5株(Buchheim et al.1997)及びChloromonas属と同定されている1株(Cr.insignis N447)のCO2濃縮機構を調査し、ピレノイドのない株との比較を行った。透過型電顕による観察で、Cd.mutabilis U578、Cd.radiata U966、Cd.augustae U1969、Cd.macrostellata S72.81には、ピレノイドスターチに囲まれた典型的なピレノイドが認められた(図4)。Cr.insignisN447では葉緑体包膜の内側に、スターチを欠くピレノイド基質様の構造が認められ(図3)、その構造にRubiscoの集中が認められた。Cd.bipapillata S11-47では、葉緑体の中央に大きな基質構造が認められたが(図2)、その電子密度は葉緑体の他のストロマ部分と変わらず、免疫電顕でRubiscoの密度も葉緑体の他の部位と変わらないことが判明したため、この構造はピレノイドでないと考えられた。これら6株ではいずれもK0.5(CO2)が10 M程度以下で、CO2に対する細胞の親和性の高さからCO2濃縮機構があることが示唆された(表2)。また、典型的なピレノイドを持つ4株のうち、Cd.mutabilis U578とCd.radiata U966では、Cd.reinhardtiiと同程度の比較的大きな細胞内無機炭素の蓄積がCO2濃縮機構の存在を支持した(表2)。一方、典型的なピレノイドを持たないCr.insignis N447とCd.bipapillata S11-47ではCO2に対する親和性とCA活性のCO2条件による変化がCO2濃縮機構の存在を強く支持した(表2)。ただし、細胞内無機炭素の蓄積は、前章で明らかになったピレノイドのないCO2濃縮機構を持つChloromonas2株と同様、Cd.reinhardtiiなどの約1/10であった(表2)。これらと前章の結果を照らし合わせると(表3)、発達したスターチに囲まれた、Rubiscoの集中した典型的なピレノイドの存在が、無機炭素の蓄積の大きさに関係していると推測された。 M程度以下で、CO2に対する細胞の親和性の高さからCO2濃縮機構があることが示唆された(表2)。また、典型的なピレノイドを持つ4株のうち、Cd.mutabilis U578とCd.radiata U966では、Cd.reinhardtiiと同程度の比較的大きな細胞内無機炭素の蓄積がCO2濃縮機構の存在を支持した(表2)。一方、典型的なピレノイドを持たないCr.insignis N447とCd.bipapillata S11-47ではCO2に対する親和性とCA活性のCO2条件による変化がCO2濃縮機構の存在を強く支持した(表2)。ただし、細胞内無機炭素の蓄積は、前章で明らかになったピレノイドのないCO2濃縮機構を持つChloromonas2株と同様、Cd.reinhardtiiなどの約1/10であった(表2)。これらと前章の結果を照らし合わせると(表3)、発達したスターチに囲まれた、Rubiscoの集中した典型的なピレノイドの存在が、無機炭素の蓄積の大きさに関係していると推測された。 4.Chloromonas属の系統解析 18S rRNAによる系統解析(Buchheim et al.1997)は、本研究で用いているピレノイドを欠くChloromonas3株とChlamydomonasの5株が近縁な単系統群であると示唆しているが、その中の系統関係は不明瞭な点が多い。今回、より詳細な系統関係を得るために調査したすべての株についてのrbcL遺伝子1128塩基対を得て系統解析を行った。その結果、最節約法と近隣結合法で同じ樹型の遺伝子系統樹が得られ、18S rRNAの遺伝子系統樹とも矛盾がなかった(図5)。今回のrbcL遺伝子系統樹は、Chloromonasとそれに近縁なChlamydomonasが2つの単系統群を形成していることを示唆した。一方はCd.mutabilis U578、Cd.radiata U966、Cd.bipapillata S11-47を含み(group A)、もう一方は残る8株から成っていた(group B)。従って、両群でピレノイドの変形・消失と、それに平行した無機炭素蓄積機能の低下が起こったことが推定された(図5)。さらにgroup Bでは、ピレノイドの消失後、CO2濃縮機構が消失したことが推定された。また、ブートストラップ値があまり高くないが、今回解析された樹形に従えば、group Bの中ではCO2濃縮機構を保持したままピレノイドの再出現が起こった可能性も示唆されうる。また、典型的でないピレノイドを持つCr.insignis N447はgroup Bの最基部に位置しているため、この株はピレノイド消失への移行型である可能性が示唆された。 <まとめ> 本研究では、自由生活性緑藻Chloromonas及びそれに近縁なChlamydomonasを用いた微細形態及び生理機能の比較生物学的調査を実施することで、i)ピレノイド(Rubiscoの集中)のない藻類においてもCO2濃縮機構は機能し得る、ii)スターチに囲まれた典型的なピレノイドの存在が、CO2濃縮機構の細胞内無機炭素の蓄積に関与している可能性がある、というCO2濃縮機構とピレノイドの機能に関する新しい示唆を得ることができた。また、本研究では用いた生物の系統解析を実施することにより、ピレノイドに関する形態と機能の両面の具体的な進化過程が明らかにされた。さらに、この系統解析の結果、i)ピレノイドのないものとそれに近縁なピレノイドを持つものの比較検討が可能となり、ii)2つの系統群で平行して、ピレノイドの変形または消失という形態進化と、それに共役した無機炭素の蓄積の低下というCO2濃縮機構の機能進化を解析することができたため、より説得力のあるピレノイドに関する機能の推測が可能となった。 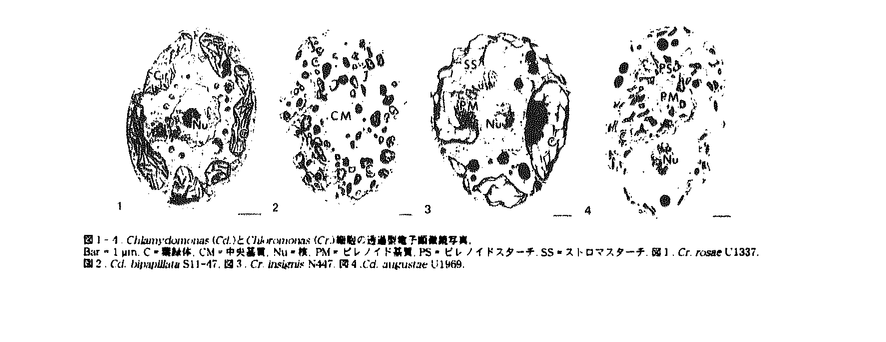 図1-4 Chlamydomonas(Cd.)とChloromonas(Cr.)細胞の透過型電子顕微鏡写真.Bar=1m.C=葉緑体,CM=中央基質,Nu=核,PM=ピレノイド基質,PS=ピレノイドスターチ,SS=ストロマスターチ.図1.Cr.rosae U1337.図2.Cd.bipapillata S11-47.図3.Cr.insignis N447.図4.Cd.augustae U1969. 図1-4 Chlamydomonas(Cd.)とChloromonas(Cr.)細胞の透過型電子顕微鏡写真.Bar=1m.C=葉緑体,CM=中央基質,Nu=核,PM=ピレノイド基質,PS=ピレノイドスターチ,SS=ストロマスターチ.図1.Cr.rosae U1337.図2.Cd.bipapillata S11-47.図3.Cr.insignis N447.図4.Cd.augustae U1969. 表1.ピレノイドのないChloromonas(Cr.)株,及びChlamydomonas(Cd)reinhardtiiにおける細胞のCO2に対する親和性(K5(CO2)).CA活性,及び細胞内無機炭素の蓄積(無機炭素濃度).細胞は空気(L)もしくは5%CO2(H)を通気して生育させたKo5(CO2)値とCA活性は平均値±SD無機炭素プールは10,20または40秒間光合成を行わせた後に測定した細胞内無機炭素濃度の最大値. 表1.ピレノイドのないChloromonas(Cr.)株,及びChlamydomonas(Cd)reinhardtiiにおける細胞のCO2に対する親和性(K5(CO2)).CA活性,及び細胞内無機炭素の蓄積(無機炭素濃度).細胞は空気(L)もしくは5%CO2(H)を通気して生育させたKo5(CO2)値とCA活性は平均値±SD無機炭素プールは10,20または40秒間光合成を行わせた後に測定した細胞内無機炭素濃度の最大値. 表2.ピレノイドのないChloromonas(Cr.)に近線なChlamydomonas(Cd)株.及びCrinsignisにおける細胞のCO2に対する親和性(Ko5(CO2)).CA活性,及び細胞内無機炭素の蓄積(無機炭素濃度).細胞は空気(L)もしくは5%CO2(H)を通気して生育させたKo5(CO2)値とCA活性は平均値±SD無機炭素プールは10,20または40秒間光合成を行わせた後に測定した細胞内無機炭素濃度の最大値. 表2.ピレノイドのないChloromonas(Cr.)に近線なChlamydomonas(Cd)株.及びCrinsignisにおける細胞のCO2に対する親和性(Ko5(CO2)).CA活性,及び細胞内無機炭素の蓄積(無機炭素濃度).細胞は空気(L)もしくは5%CO2(H)を通気して生育させたKo5(CO2)値とCA活性は平均値±SD無機炭素プールは10,20または40秒間光合成を行わせた後に測定した細胞内無機炭素濃度の最大値. 表3.2と3のまとめ. 表3.2と3のまとめ. 図5.最節約法で構築したrbcL遺伝子系統樹.枝の数字は50%以上のブートストラップ値を示す.<引用文献>Buchheim et al.(1997)J Phycol 33:286.Palmqvist et al.(1994)Planta 195:70.Suss et al.(1995)Plant Physiol 107:1387. 図5.最節約法で構築したrbcL遺伝子系統樹.枝の数字は50%以上のブートストラップ値を示す.<引用文献>Buchheim et al.(1997)J Phycol 33:286.Palmqvist et al.(1994)Planta 195:70.Suss et al.(1995)Plant Physiol 107:1387. |
| 審査要旨 | | 本論文は4章からなり、第1章は、ピレノイドにおけるRubiscoの集中、第2章は、ピレノイドを欠くChloromonas株の形態及びCO2濃縮機構、第3章は、Chloromonasに近縁なChlamydomonas等の株の形態及びCO2濃縮機構、第4章はChloromonas属及びそれに近縁なChlamydomonas株の系統解析について述べられている。光合成CO2固定酵素のRubiscoは、通常の陸上植物では葉緑体ストロマ中に分散して存在しているが、多くの真核藻類の葉緑体内では集合して基質構造をとり、スターチに囲まれ『ピレノイド』として存在している。しかしながら、ピレノイドの生物学的意義については不明な点が多く、その生理的役割を直接的に証明した研究例はない。藻類では、細胞としてのCO2に対する親和性は高くなっており、この現象はCO2をRubiscoの回りに集める『CO2濃縮機構』が働いていることで説明されている。一方、藻類の中にはピレノイドを持たない種もあり、近年、地衣類の共生藻を用いた比較生理学的研究が行われ、ピレノイド構造の生理的役割が議論された(e.g.Palmqvist et al.1994)。これらの研究では、用いたピレノイドを欠く藻類すべてがCO2濃縮機構を欠いており、ピレノイドがCO2濃縮機構に必須であると議論された。しかしここで調査された藻類は地衣類の共生藻に限られており、近縁なピレノイドを持つ種との比較もなされていない。本論文では、真核藻類で最もCO2濃縮機構の研究の進んだ種の一つである単細胞緑藻Chlamydomonas(Cd.)reinhardtiiが所属するChlamydomonas属に近縁でかつピレノイドを欠くとされる自由生活藻Chloromonas(Cr.)属、並びにChlamydomonas属を材料とし、その葉緑体のピレノイドに関する微細形態とCO2濃縮機構を比較し、ピレノイドの機能を推測することを目的とした。第1章は、本論文の研究開始直後発表されたSuss et al.(1995)の研究に関係しており、Suss et al.(1995)は凍結固定法を用いた免疫電子顕微鏡(電顕)法による観察結果から、Cd.reinhardtiiのRubiscoの大部分はピレノイドではなくストロマに存在していると報告した。これはピレノイドがRubiscoの集中した構造である、という本論文の前提と矛盾するため、Cd.reinhardtiiのRubisco抗体を用いて再検査した。その結果、凍結法を用いても従来の常温での固定法と同様の結果が得られ、Cd.reinhardtii細胞でRubiscoがピレノイドに集中していることが再確認された。第2章は、Chloromonas株の中で、光学顕微鏡と透過型電顕でピレノイド構造のないことが確認され、さらに免疫電顕で、葉緑体内にRubiscoの集中した部位が認められなかった、5株でCO2濃縮機構の検討を行った。その結果、2株でCO2濃縮機構を保持していることがピレノイドがない藻類で初めて明らかになった。第3章では2章で調査したピレノイドのないChloromonas属に近縁とされるChlamydomonas5株及びChloromonas属と同定されている1株(Cr.insignis N447)の葉緑体の微細構造、Rubiscoの分布及びCO2濃縮機構を調査した。これらの中で4株は典型的なピレノイドを持ち、CO2濃縮機構の存在が明かになった。一方、典型的なピレノイドを持たない他の2株ではCO2濃縮機構の存在が示唆されたが、細胞内無機炭素の蓄積は、2章で明らかになったピレノイドのないCO2濃縮機構を持つChloromonas2株と同様、Cd.reinhardtii等の約1/10であった。したがって、スターチに囲まれた、Rubiscoの集中した典型的なピレノイドの存在が、無機炭素の蓄積の大きさに関係していると推測された。第4章は詳細な系統関係を得るために調査したすべての株についてのrbcL遺伝子1128塩基対を得て系統解析を実施した。その結果、Chloromonasとそれに近縁なChlamydomonasが2つの単系統群を形成していることが示唆され、両群でピレノイドの変形・消失と、それに平行した無機炭素蓄積機能の低下が起こったことが推定された。更に片方の群の中でピレノイドの消失後、CO2濃縮機構が消失したことが推定された。 以上の様に本論文では今までに光合成の研究分野で用いられていなかった新しい材料を用いることで、細胞構造に関する新しい機能推測を可能とした点で大きく植物細胞学に寄与している。また、細胞の微細構造と生理機能の両者の進化過程を分子系統学的解析に基づき議論するという今まで植物系統学分野では見られなかった新たなる研究分野を切り開いた点も大いに評価されうる。なお、本論文第1章は黒岩晴子氏その他2名、第2章は阿部俊彦氏その他6名との共同研究であるが、論文提出者が主体となって実験、観察及び検証を行ったもので、論文提出者の寄与が十分であると判断する。 したがって、博士(理学)の学位を授与できると認める。 |