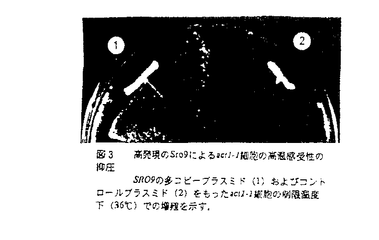
本学位論文は出芽酵母を実験材料として細胞極性の確立/維持の分子機構を研究したものである。本研究分野に関する研究の現状を概説した後、第1章では新規遺伝子SRO9の機能について、第2章ではもう一つの新規遺伝子SRO7の機能解析について研究し、その結果から、アクチン細胞骨格系形成の制御による細極性の確立/維持機構に関するモデルを提案している。以下にその概略を述べる。 序論 細胞が機能を果たし、生育するためには、細胞内での物質分布等の不均一性が必要である。このような非対称性の形成には、細胞の極性の存在が必須である。細胞極性の存在は、神経細胞の軸索成長、神経伝達物質の分泌、発生期における分化の決定、など高等生物から、単細胞真核生物である酵母の出芽、接合に至るまで、多様な細胞で多くの生命現象に見られる。細胞極性がどのようにして確立し、維持されていくかの分子レベルでの解析は、まだよくなされていない。出芽酵母における細胞極性は、出芽過程を通じて観察される。酵母は、出芽時に、アクチン細胞骨格が非対称的に分布し(アクチンパッチは、細胞が成長している部位に分布し、アクチンケーブルは細胞の長軸方向に沿って伸びる)細胞壁や細胞膜の成分を成長部位へ運ぶと考えられている。細胞の形態変化にはrho型GTPaseが関与していることが報告されている。酵母には6種のrho型GTPaseが存在する(Cdc42、Rho1、2、3、4、5)。Cdc42、Rho1は細胞増殖に必須であり、Rho3は30℃以上で必須である。Rho3は、その機能を失うと細胞は小さな芽を出したところで細胞溶解し、極性を失った表現型を示すことから、芽の成長に必要である。Rho3の機能欠損を高発現で抑圧する遺伝子(SRO遺伝子)としてRHO4、BEM1(SRO1)、CDC42(SRO2)、SEC4(SRO6)、SRO3、4、5、7、8、9が単離されている。Bem1は出芽及び接合に必要な因子でアクチンと結合する。Sec4は輸送小胞の細胞膜への融合に必要な因子である。従って、Rho3はCdc42、Rho4と共同して、アクチン細胞骨格の方向を制御することで、芽の成長を制御していると考えられる。Rho3の機能欠損を多コピーで抑圧するSRO遺伝子群の解析は、極性に従った、膜成長や細胞骨格系の構築の機構の解明に重要である。本研究において、SRO遺伝子群、特にそのうちのSro7、Sro9の機能解析を通して、細胞極性維持機構の遺伝学的、分子生物学的解析が行われた。 第1章トロポミオシンと類似機能を持つSro9の解析 SRO9遺伝子は466アミノ酸からなる親水性蛋白質をコードする。Sro9蛋白質の細胞内での機能を調べるためにSRO9遺伝子破壊株を作成したが、その株は、野生型と変わらない表現型を示した。芽の成長には、厳密に制御されたアクチン細胞骨格系の再構成や、それにともなった、膜成分などの輸送が必要であり、また、他のSRO遺伝子も、この過程に関わっているものがあることから、アクチン細胞骨格系の遺伝子との遺伝学的関係を調べた。その結果、アクチン細胞骨格系の因子で、アクチンファイバーに結合して、アクチンケーブルを安定化することで物質輸送に関与するトロポミオシンと類似機能をもつことが明らかとなった。すなわち、SRO9はトロポミオシン遺伝子TPM1の欠損と合成致死性を示す。sro9 tpm1二重変異株細胞はアクチンケーブルの消失、アクチンパッチの非局在化など、極性を失ったときに典型的にみられる表現型を示し細胞溶解する。tpm1 細胞では細胞中のアクチンケーブルが消失し増殖が遅延するが、SRO9は高発現によりtpm1遺伝子破壊株の増殖遅延を細胞中のアクチンケーブルを回復することにより抑圧できること。rho3遺伝子破壊株は、細胞極性を失いアクチンが細胞全体に非局在化し増殖阻害が生じるが、TPM1もSRO9と同様に高発現により、その細胞中のアクチンケーブルを回復することで、rho3遺伝子破壊株の増殖阻害を抑圧した。 細胞では細胞中のアクチンケーブルが消失し増殖が遅延するが、SRO9は高発現によりtpm1遺伝子破壊株の増殖遅延を細胞中のアクチンケーブルを回復することにより抑圧できること。rho3遺伝子破壊株は、細胞極性を失いアクチンが細胞全体に非局在化し増殖阻害が生じるが、TPM1もSRO9と同様に高発現により、その細胞中のアクチンケーブルを回復することで、rho3遺伝子破壊株の増殖阻害を抑圧した。 Sro9のアクチン細胞骨格系への関連が示されたので、アクチンそれ自身との遺伝学的関係を調べた。出芽酵母のアクチンは必須遺伝子ACT1によってコードされる。ACT1には多数の変異が同定されているが、そのうちの一つであるact1-1は、温度感受性を示し、許容温度下でも細胞中のアクチンが細胞全体に非局在化する。この温度感受性を、高発現のSRO9はアクチンケーブルは回復できないものの、アクチンパッチの出芽部位への局在を回復することで、部分的に抑圧できた。一方、TPM1は、増殖もアクチンの局在も回復できなかった。 さらに、sro9tpm1二重遺伝子破壊株の細胞溶解の表現型を指標として関連遺伝子の検索を行った。その結果、もう一つのトロポミオシン遺伝子TPM2が単離された。TPM2単独の遺伝子破壊は、特に表現型を示さないが、tpm1tpm2二重遺伝子破壊株は、増殖できなくなる。このトロポミオシン完全欠失株の致死性を、tpm1単独のときと同様その増殖遅延をを抑圧できるかを調べたが、SRO9はこの致死性は回復できなかった。また、Sro9pの細胞内局在を同定したところ、トロポミオシンとは異なり、細胞質中に存在することがわかった。 これらのことから、Sro9の細胞内での機能は、それ自身単独ではアクチンファイバーの安定化能はもたないが、トロポミオシンと同様に、適切な物質輸送が行えるように、アクチンが正しく出芽部位に局在するように働くことで、芽が正常に成長するよう機能していると考えられる。 第2章細胞質分離に関与するSro7の解析 SRO7遺伝子は1033アミノ酸からなる蛋白質をコードする。データベースの検索から、出芽酵母にはもう一つSro7pとよく似た蛋白質(55%identical)をコードするSRO77の存在がわかった。各々単独の遺伝子破壊は増殖に影響を与えないが、二重遺伝子破壊株sro7sro77は、20℃以下で低温感受性を示した。そのとき細胞は、わずかに丸く大きな形になり、アクチンが部分的に非局在化するなど細胞極性の維持に欠陥を生じた。また、Factin形成阻害能を持つ薬剤、Latrunculin-Aにより酵母細胞のアクチン細胞骨格形成は影響をうけるが、sro7sro77二重遺伝子破壊株は、Latrunculin-A超感受性を示し、アクチン細胞骨格系への関与が示唆された。これらのことから、Sro7、Sro77の細胞内での機能は、アクチン細胞骨格系を極性に従って、配向させるよう働いていると考えられる。 他の生物種のアミノ酸配列との比較により、Sro7/Sro77は、Dorosophilaのガン抑制遺伝子lethal(2)giant larvae[l(2)gl]とよく似たドメインをもつことがわかった。l(2)glが機能欠損したDorosophilaは、発生過程で分化に必要な極性が規定されず、腫瘍を形成してしまう。ヒトのl(2)glのカウンターパートLLGLをsro7sro77二重遺伝子破壊株導入したところ、低温感受性をを相補したことことから、Sro7/Sro77はl(2)glの酵母のカウンターパートと考えられる。l(2)glはType II Myosinと結合することが報告されている。そこで、酵母のType II MyosinであるMyo1とSro7との結合を免疫沈降により調べたところ両者は共沈した。さらに、GST-Sro7結合GSH beadsとMyo1が共沈した)ことから、その結合が確認された。従って、Sro7とMyo1は、細胞内で複合体を作っていると考えられる。 次に、間接蛍光抗体法によりSro7、Myo1の局在を調べたところ、Myo1はbud neckに、Sro7は細胞表層に存在していた。Sro7は細胞分画法により調べると、細胞膜及び、細胞質両方に存在していた。この局在は、l(2)glの局在と同じである。 両者の関係をより調べるために、遺伝学的解析をおこなった。その結果、Sro7、Sro77とMyo1は、相互の機能欠損による増殖阻害を各々の遺伝子破壊により抑圧しあうことがわかった。すなわち、MYO1は、遺伝子破壊により細胞質分裂に欠損を生じ、増殖が遅くなるが、sro7sro77myo1三重遺伝子破壊株は、野生型と同等の増殖速度を示し、細胞質分裂もほぼ正常であることを明らかとした。従って、Sro7、Sro77は、Myo1が適切に細胞質分裂を行えるように制御していると考えられる。 Myo1や、Myo2などの細胞内の運動性因子を本来の経路のみで働くように制御することにより、正常なアクチン細胞骨格系の機能を維持させていると考えている。 以上のように、本研究により出芽酵母の芽の成長過程に関わっている因子が、アクチン細胞骨格系と機能的に連関していることが明らかになった。極性成長の一つのモデルである出芽過程で、その初期段階であるアクチン細胞骨格系の再構成から、最終段階である細胞質分裂まで働く因子群を、分子スイッチとして働く低分子量GTPaseが制御していることを明らかとしたことで、本研究は極性成長という普遍的な現象の分子レベルでの解明への端緒を開いたといえる。発表論文は共著であるが、SRO7とSRO9遺伝子の機能解析は論文提出者独力によって行われたものであることが認められた。このような考慮から、本論文は博士(理学)の学位に値するものであることが審査員全員の一致により認められた。 | 

 mを示す.
mを示す.