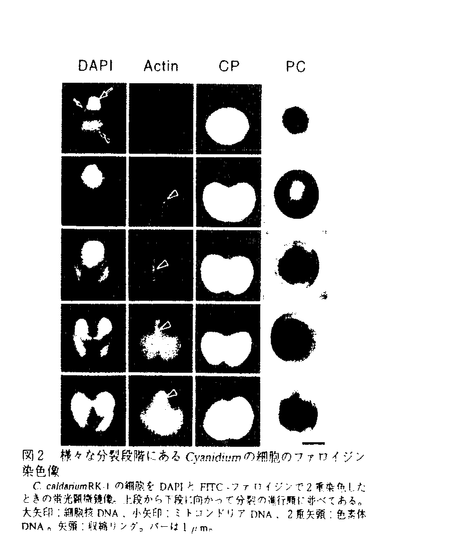
本論文は4章からなり、第1章はCyanidiumのアクチン遺伝子の単離とその解析について、第2章・第3章はそれぞれCyanidiumの細胞の分裂様式の蛍光顕微鏡観察と電子顕微鏡観察について、第4章はCyanidioschyzonの分裂様式の解析と真核生物に見られる細胞の分裂装置の進化について述べられている。 細胞の分裂という現象は最も基本的かつ重要な生命現象の1つであり、真核生物ではアクチンからなる収縮リングの収縮により引き起こされる。従来の主たる研究材料は動物卵であるが、その収縮リングは巨大かつ複雑であるため、収縮時の微細構造の変化や収縮力の発生機構の解析は困難であった。そこで、論文提出者は原始紅藻のCyanidiumとCyanidioschyzonを研究材料に選んだ。両者は動物卵の僅か100分の1の大きさにすぎない。このうちCyanidiumでは2層の極めて単純な収縮リングが観察されるため、従来の研究では欠けていたアクチン遺伝子の発現から収縮リングの構築・収縮までの一連の過程を解析するのに適している。一方、Cyanidioschyzonでは収縮リングが観察されないため、両者の分裂様式を比較・解析することで真核生物の分裂装置の進化を明らかにできる。論文提出者はこうした観点から、細胞質分裂の基本原理と分裂装置の進化の解明を目標として研究を行った。 遺伝子発現の流れに沿って、第1章ではCyanidiumのアクチン遺伝子を単離・解析した結果、(1)アクチン遺伝子は2つ存在しているが377個の同一のアミノ酸配列をもつタンパク質をコードしており、(2)それらは同一の17番染色体上に存在することを明らかにした。そして、(3)他の真核生物のアクチンmRNAと同じ1.5kbの大きさで転写されていることも示した。 第2章では、(1)Cyanidiumの細胞質分裂の同調系を開発し、(2)同調化された細胞のファロイジン染色を行った結果、分裂に先立って形成された収縮リングはアクチン繊維量を一定に保ったまま収縮して、徐々に太くなることを明らかにした。さらに、(3)Cyanidiumのアクチンに対する抗体を作成して細胞の抗体染色を行い、収縮リングはG-アクチンを含むドットからde novoに形成されることを明らかにした。 第3章では、Cyanidiumの分裂様式を電子顕微鏡により詳細に解析している。その結果、(1)収縮リングの収縮過程で、内層・外層ともに厚さは不変だが幅を増加させながら滑り込むことを明らかにした。また、(2)アクチンドットは小胞の形をとっていることも明らかにした。そして、(3)細胞をアセトンのみで固定することにより、免疫電子顕微鏡法において従来の手法の2.6倍の検出感度が得られることを発見した。(4)この新手法で固定した細胞に対して抗アクチン抗体処理した結果、収縮リング2層のうちの内層がアクチン繊維であることを示した。このことから論文提出者は、外層がミオシンであり、内層のアクチンとの間で滑り込みを起こして収縮力を発生するモデルを提起している。 Cyanidiumとは近縁であっても、Cyanidioschyzonには収縮リングは観察されない。そこで、第4章ではCyanidioschyzonのアクチンに対する抗体を作成し、(1)ウエスタンブロッティング、(2)間接蛍光抗体染色、(3)免疫電子顕微鏡法による解析を行っている。いずれにおいても反応が見られなかったことから、Cyanidioschyzonは原始的であるがゆえに極めて少量のアクチンにより細胞質分裂が引き起こされている可能性が示唆された。そこで論文提出者は、(4)アクチン遺伝子の塩基配列をもとに系統樹を作成してCyanidioschyzonの原始性を証明するとともに、(5)系統樹をもとに真核生物で見られる細胞の分裂装置の進化を推察している。まず最初にCyanidioschyzonで見られるような極めて少量のアクチンからなる分裂装置が登場し、次いでCyanidiumの収縮リングのような20数本のアクチンからなる分裂装置へと進化し、その後のさらなる進化の過程で分裂装置を構成するアクチン繊維の数は増大し、分裂装置自身も巨大化・複雑化・多様化していったと考えている。 論文提出者は、「細胞の分裂」という生物における基本的問題を解明するためにCyanidiumという、これまで使われていないが研究を遂行する上で有利な生物に着目し、遺伝子発現の流れに沿って、分子生物学的手法と形態観察により細胞質分裂の機構を総合的に解析している。このような研究は皆無である。さらには、推測の域にすぎなかった収縮リングの滑り込みの過程を初めて経時的にとらえて定量化した研究は注目されている。一方、Cyanidioschyzonの分裂様式の解析により、真核生物の分裂装置の進化を明らかにした研究も独創的である。 なお、本論文第1章は、高野博嘉・伊藤竜一・河野重行・黒岩常祥と、第2章は、高野博嘉・黒岩晴子・戸田恭子・黒岩常祥と、第3章は、黒岩晴子・宮城島進也・伊藤竜一・戸田恭子・黒岩常祥と、第4章は、黒岩晴子・宮城島進也・伊藤竜一・戸田恭子・黒岩常祥との共同研究であるが、論文提出者が主体となって分析および検証を行ったもので、論文提出者の寄与が十分であると判断する。従って、博士(理学)の学位を授与できると認める。 |  mと巨大で、収縮リングも幅が広く多数のアクチン繊維が入り組んでいる。そのため、収縮リングの微細構造や収縮力の発生機構などの解析は困難で、生化学的な研究はある程度なされているものの、細胞質分裂の分子細胞学的研究は進展していない。これに対して、より原始的で極めて小さな生物ならば収縮リングも小さくて単純な構造であると予測されることから、細胞質分裂の機構の研究に関しても進展が期待できる。そこで、原始紅藻のCyanidium caldarium RK-1を研究材料に選んだ。Cyanidiumは直径わずか数
mと巨大で、収縮リングも幅が広く多数のアクチン繊維が入り組んでいる。そのため、収縮リングの微細構造や収縮力の発生機構などの解析は困難で、生化学的な研究はある程度なされているものの、細胞質分裂の分子細胞学的研究は進展していない。これに対して、より原始的で極めて小さな生物ならば収縮リングも小さくて単純な構造であると予測されることから、細胞質分裂の機構の研究に関しても進展が期待できる。そこで、原始紅藻のCyanidium caldarium RK-1を研究材料に選んだ。Cyanidiumは直径わずか数