1961年、Nirenbergらが大腸菌抽出液に鋳型mRNAや、アミノ酸、エネルギー物質などを添加した無細胞タンパク質合成系を用いて遺伝暗号の研究を発表した。以後、小麦胚芽、ウサギ網状赤血球などからの抽出液を用いる多くの無細胞タンパク質合成系が開発された。さらに、DNAからmRNAを転写しながらタンパク質を合成する転写翻訳共役系も開発された。これらの無細胞タンパク質合成系は遺伝学的或いは生化学的研究の手段として広く利用されてきている。 従来、無細胞タンパク質合成はバッチ方式で行われてきたが、タンパク質合成反応は1-2時間で停止する。得られるタンパク質量がかなり少ないため、多くの場合、タンパク質の検出は放射性同位元素や免疫学的な手段でしかできない。すなわちタンパク質の生産手段としては利用できなかった。また、このことは遺伝子の転写及び翻訳等の研究にも大変不便を与えている。 1988年、Spirinらは連続方式で高効率のタンパク質合成に成功した。細胞抽出液とmRNAを板状濾過膜反応器へ充填し、タンパク質合成に必要な成分、ATP、GTP、アミノ酸を連続的に供給することで、タンパク質合成反応が数10時間継続するという現象を見いだした。そしてこの条件では従来より大量にタンパク質を得ることができた。しかしこの技術は再現が難しく、追試も難航しており、現在限られたグループしか追試に成功していない。さらに、Spirinらの方法では実験を行う際に、二つの恒温器、ポンプ、膜を装着した反応器、コレクター、タンク、管などの複雑な装置を必要とする 本研究では従来のバッチ式無細胞タンパク質合成反応の主な停止原因を究明することを第一の目的とした。さらに、その停止原因を解消し、簡便かつ高い合成能力を有する無細胞タンパク質合成法の開発を第二の目的とした。本研究では、まず小麦胚芽抽出液を用いる従来のバッチ式無細胞タンパク質合成系でタンパク質合成反応の主な停止原因を分析した。その結果、タンパク質合成反応の主な停止原因はエネルギーチャージ((│ATP│+│ADP│/2)/(│ATP│+│ADP│+│AMP│))の低下であることが分かった。また、エネルギーチャージの低下はタンパク質合成の有無と関係なく、主に抽出液由来の酸性phosphataseによることを見出した。タンパク質合成速度はエネルギーチャージと深く関係する。エネルギーチャージが低下すると、タンパク質合成速度は急激に下がる。特にエネルギーチャージが0.85以下に下がると、タンパク質合成はほぼ停止する。そこで我々はエネルギーチャージを0.85以上に戻すことによって、タンパク質合成が再開するか否か、検討した。すなわち30℃でタンパク質合成を1時間行った反応液を(1)そのままさらに1時間incubationした場合と(2)遠心濾過でATP、GTPの加水分解物などの低分子成分を除去した後、ATP、GTP等を再添加するエネルギー更新を行い、エネルギーチャージを高めた後、再度30℃でタンパク質合成を1時間行った場合のタンパク質合成能力を比較した。その結果、エネルギー更新を行った場合にはタンパク質合成が再開することを確認した(図1)。さらに、このエネルギー更新を行った後の1時間で得られたタンパク質量が、最初の1時間のincubationで得られたタンパク質合成量より多いことを見い出した。このことから、最初の1時間のincubation過程、或いはエネルギー更新過程で、何らかの原因でタンパク質合成能力が増加するのではないかと考えた。 次に、タンパク質合成能力の増加方法について検討を行った。従来のバッチ式無細胞タンパク質合成系では、30℃で合成したluciferaseタンパク質がincubation時間20分以降から検出された。タンパク質合成量の増加は1時間で停止することから、30℃でのタンパク質合成反応の継続時間は実質的には40分前後であることが分かった。そこで、タンパク質合成能力の増加には1)合成開始誘導時間を短くする;2)タンパク質合成速度を速くする;3)エネルギー更新によりタンパク質合成を継続させることが有効と考えた。  図1 エネルギー更新の効果。エネルギー更新でエネルギーチャージが回復、タンパク質合成が再開 図1 エネルギー更新の効果。エネルギー更新でエネルギーチャージが回復、タンパク質合成が再開 タンパク質合成反応は開始、延長、停止の三段階から構成されている。開始過程はアミノ酸とtRNAのaminoacylationから始まり、三重複合体、43S、48S、80S開始複合体の形成を含んでいる。そのうち、48S開始複合体の形成がタンパク質合成反応の律速段階であることがこれまでの研究で分かっている。48S開始複合体は43S開始複合体とmRNAから成る。したがって、43S開始複合体を前もって合成することでタンパク質合成が早く始まると予想された。タンパク質合成開始誘導時間が短ければ、反応系のエネルギーチャージが低下する前に合成反応が進むので、結果的にタンパク質合成能力が増加すると考えられた。また、43S開始複合体の生成速度を高めることができれば、48S開始複合体の生成速度、すなわちタンパク質合成速度を増加させることも可能であると考えられた。 そこで、本研究ではあらかじめ43S開始複合体を形成させた後、エネルギー更新でエネルギーチャージを高めて、タンパク質合成を行う新しいタンパク質合成方法を考案し、これをPER法(preincubation followed by energy replenishment method)と名付けた(図2)。まず、小麦胚芽抽出液とATP、GTP、CP(creatine phosphate)、アミノ酸などを含むincubation液をpreincubationし、43S開始複合体の形成を行う。この過程ではATP、GTPが加水分解されるので、遠心濾過チューブを用いて、incubation液を遠心濾過し、ATP、GTPの加水分解物等の低分子成分を除く。低分子成分を除去したincubation液に新しいATP、GTP、CP、アミノ酸などを含むreplenishment bufferを濾過液と等量加え、さらに鋳型mRNAを加えて30℃でタンパク質合成を行う。この操作方法により、従来のバッチ式無細胞タンパク質合成と較べ最高10倍のタンパク質合成能力が得られた(図3及び図4)。PER法が高いタンパク質合成能力を有する原因として(1)preincubationによる43S開始複合体の形成;(2)エネルギー更新によるエネルギーチャージの回復;(3)遠心濾過による43S開始複合体の形成の促進などが考えられる。これら3つの考えうる原因のタンパク質合成能力増加への寄与を明らかにするための実験を行った。その結果、以下のことが判明した。Preincubation及びエネルギー更新はタンパク質合成能力増加に大いに貢献する。さらに、preincubation時におけるエネルギー物質の存在がタンパク質合成能力の増加に特に大きい効果を与える。すなわち、preincubation過程でエネルギーを要する三重複合体や43S開始複合体などが予め生成するため、タンパク質合成反応が早く開始し、タンパク質合成速度が増大し、タンパク質合成能力が大幅に増加する。また、エネルギー更新過程ではincubation液が一旦濃縮されるため、三重複合体や43S開始複合体などの生成がさらに促進され、タンパク質合成能力が増加すると考えられる。 PER法は簡便且つ高効率な無細胞タンパク質合成法としてタンパク質生産に適用できる。また、エネルギー更新が加えられているため、エネルギーチャージ低下の影響を排除しつつ、タンパク質合成機構の研究および鋳型mRNAの分解や第三組成の添加のタンパク質合成への影響の研究ができる。さらに、PER法は遠心濾過チューブを反応器とする回分法のため、複雑なシステムを必要とする連続無細胞タンパク質合成法(Spirinらの方法)よりかなり手軽にエネルギー更新、途中での任意成分の追加、反応器内の反応液のサンプリングなどができる。さらに、PER法で遠心濾過チューブのかわりに膜型反応器を用いることにより工業的規模でのタンパク質生産を周期的に行うシステムの実現が可能になると考えられる。  図2 PER(preincubation followed by energy replenishment)法概念図。 図2 PER(preincubation followed by energy replenishment)法概念図。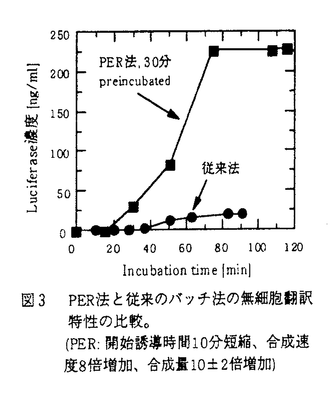 図3 PER法と従来のバッチ法の無細胞翻訳特性の比較。PER:開始誘導時間10分短縮、合成速度8倍増加、合成量10±2倍増加 図3 PER法と従来のバッチ法の無細胞翻訳特性の比較。PER:開始誘導時間10分短縮、合成速度8倍増加、合成量10±2倍増加 図4 PER法におけるpreincubation時間の効果。バッチ法のタンパク質合成量を1としたPER法の合成量 図4 PER法におけるpreincubation時間の効果。バッチ法のタンパク質合成量を1としたPER法の合成量 |