自然突然変異は、生物が通常の環境下で生育するときにごく希に生じる突然変異のことである。自然突然変異は進化の原動力となるばかりでなく、今日では老化や発ガンのプロセスに密接に関与した現象であることが知られている。自然突然変異の研究は主として大腸菌を材料として進められてきた。これは自然突然変異が極めて希な現象であるため研究に大量の細胞集団が必要であったことと、大腸菌では古くからミューテーター変異株と呼ばれる突然変異を高頻度におこす変異株の存在が知られていたことによる。これまでのミューテーター変異株をもちいた遺伝学的な研究から自然突然変異の主要な原因は、DNA複製での複製エラーであることが明らかになった。また大腸菌は複製エラーの発生自体を抑制する機構と発生した複製エラーを修復する機構を複数有しており、これらの機構は広く生物一般に保存されていることも近年では認識されている。このような状況で我々の研究グループは、複製エラーの実体とその発生機構の研究を生化学的な手法により試みている。従来の遺伝学的手法とは異なり、精製された大腸菌野生型DNA polymerase III(Pol III)により試験管内DNA複製反応を行わせ、この時発生する複製エラーを検出、解析して複製エラーの種類や特徴、塩基配列特異性を明らかにする。つぎに各種の複製エラーについて、試験管内DNA複製反応の条件を様々に変化させることによりその発生機構を解明していく手法である。
突然変異検出のレポーター遺伝子であるrpsL遺伝子をもつoriCプラスミドDNAを鋳型として試験管内ローリングサークルDNA複製を行わせ、その時発生する複製エラーについて100サンプル、塩基配列レベルで解析した。これまでDNAポリメラーゼは主に塩基置換を起こすと考えられていたが、意外なことに検出された複製エラーの半数以上が1塩基フレームシフト変異であった。またこの変異は特にdA/dTトラクトと呼ばれるモノヌクレオチドリピート配列に集中して発生しており、その多くが1塩基付加型(+1)のものであった。dA/dTトラクトにおいて+1フレームシフト変異が高い頻度で発生することは、T4ファージをもちいたin vivoの実験で古くから知られていたがその発生機構はまだ解明されていない。また、ヒトでの突然変異の解析はがんや遺伝病の研究の進展に伴って急速に発展しており、ここ数年の間にヒト遺伝子に発生した突然変異の解析結果が急速に蓄積されてきている。それらの中にはdA/dTトラクトでの+1フレームシフト変異が多数含まれており、この変異の発生機構は医学的な面からも興味深い現象である。本研究はこのような背景に立ち、dA/dTトラクトでのフレームシフト変異がどのような機構でDNAポリメラーゼにより引き起こされるのか、その発生機構を生化学的に解明することを目的として行われた。
A.単鎖DNAを鋳型とした試験管内DNA複製系を用いた+1フレームシフト変異の研究 dA/dTトラクトでのフレームシフト変異の発生機構を生化学的に研究するには、この変異がdAトラクト、dTトラクトのどちらのトラクトを鋳型としてDNA複製を行う際に発生するのかを決定できる実験系が望ましい。これまで我々が使用してきたoriCプラスミドDNAを鋳型とする試験管内DNA複製系は、そのような実験には不向きである。またoriC実験系の場合、試験管内DNA複製に10種類以上のタンパク質が必要であり、この中には直接DNA複製には関係のないタンパク質が多く含まれている。これらのタンパク質群の中にはDNA結合活性を示すタンパク質が含まれており、それらがDNA複製に及ぼす影響や、持ち込みの塩の影響等不確定な要素が多い。そこでまずはじめにdA/dTトラクトでのフレームシフト変異を研究するための試験管内DNA複製系を新規に構築した。また同時に、コロニーハイブリダイゼーションを用いることにより大量のサンプルの中からdA/dTトラクトでの+1フレームシフト変異を特異的に検出する検出系の開発も行った。
これらの実験系を用いてフレームシフト変異の発生機構を解析した結果、この変異の発生には以下の大きな特徴があることが明らかになった。
a)この変異はdTトラクト上を複製する際に高い頻度で発生する。
b)この変異はdATP濃度に依存して発生する。
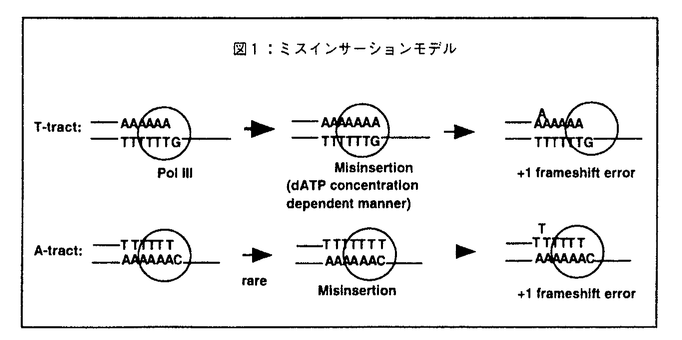
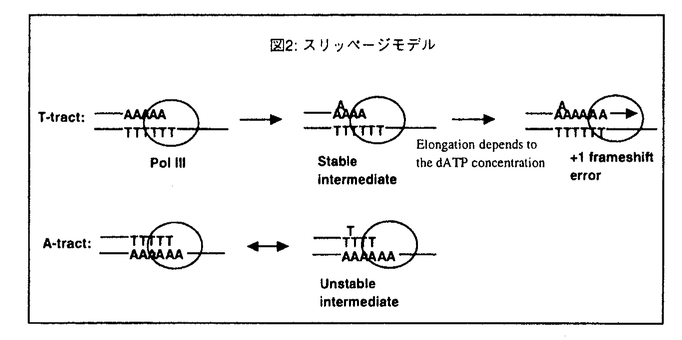
これらの実験事実のうちdATP濃度依存性については、2通りの解釈が可能である。1つは鋳型となるトラクトの5’残基に対するdATPのミスインサーションが起因となってこの変異が発生する可能性(ミスインサーションモデル)であり、今一つはトラクト上でスリッページエラーにより形成された+1フレームシフト変異中間体からの鎖伸長反応の頻度をdATP濃度が規定している可能性(スリッページモデル)である。まずはじめに前者の可能性が高いのではないかと考え以下の実験をおこなった。
(1)ミスインサーションモデルの検証 Kunkel等により提唱されたミスインサーションモデルでは、鋳型dTトラクトの5’側の残基(-dG)に対して誤った塩基対を形成した残基(dA)がずれることにより+1フレームシフト変異が発生する(図1)。このモデルに従えばdAトラクトを複製する際には鋳型の-dT残基に対するdTTPのミスインサーションが起因になることになる。しかしながら、鋳型dTに対するdTTPのミスインサーションは極めて希にしか起きないことが知られている。そのためdAトラクトを鋳型とする場合にはこの変異はほとんど発生せず、そのためdTトラクトを複製する際に主として発生することになる。またミスインサーションの頻度はdATPの濃度に比例することから、前記の特徴をすべて明確に説明するものと思われる。
DNA polymerasel(Poll)等のprocessivityの低い酵素がdA/dTトラクトをはじめ多くの塩基配列上で、このモデルにそった機構で-1塩基フレームシフト変異を起こすことはすでに明らかになっているが、このモデルがPol IIIのような極めてprocessivityの高い酵素によるdA/dTトラクトでの+1フレームシフト変異に適用できるかはまだ知られていない。そこでこのモデルに対する以下の検証実験を行った。
(a)高濃度dCTPによる+1フレームシフト変異発生の阻害効果
ミスインサーションが起因となってこの変異が発生するのであるならば、トラクトの5’側残基(-dG)に相補的なdCTPを大量に反応系に加えればミスインサーションが抑制され、劇的な阻害効果が観察されるはずである。しかしながら、高濃度dCTP存在下でも+1フレームシフト変異の発生頻度に大きな差は観察されなかった。
(b)鋳型5’残基による+1フレームシフト変異の発生頻度の変化
ミスインサーションの頻度は、鋳型となる塩基によって大きく異なる。したがって、トラクトの5’側の残基を様々に変化させればこの変異の発生頻度もミスインサーションの頻度に応じて大きく変化するはずである。そこでトラクトの5’側残基を部位特異的変異導入法により様々に変化させ、+1フレームシフト変異の発生頻度を測定した。しかしながら、ミスインサーションの頻度とこの変異の発生頻度に相関関係は観察されなかった。
以上の結果から、この変異は従来のミスインサーションモデルとは異なる機構により発生しているものと思われる。そこでdATP濃度依存性を説明するもう1つの考えであるスリッページモデル(図2)に基づいた解析をおこった。
(2)鋳型特異性の発生機構 dA/dTトラクト上でのスリッページエラーによりこの変異が発生する場合、なぜdTトラクトで高い頻度で発生するのか?という疑問が生じる。1つの可能性はdTトラクトを鋳型として複製する場合とdAトラクトを鋳型として複製する場合とではスリップの頻度が大きく異なる可能性である。今1つの可能性は、スリップにより生じた+1フレームシフト変異中間体の安定性が大きく異なる可能性である。後者の可能性について、+1フレームシフト変異中間体の安定性をオリゴマーDNAを用いた融解温度(Tm)の測定実験により比較したところ、どちらのトラクトを鋳型とした場合でも中間体の安定性に有意な差は観察されなかった。従って、スリップの頻度そのものが大きく異なる可能性が高い。今後は、+1フレームシフト変異中間体の発生頻度の違いを検証する予定である。