| 内容要旨 | | 酵母は単細胞で増殖する最も単純な真核生物であり、高等真核生物のモデルとして有用である。特に細胞分裂の制御機構は分子レベルだけでなく制御系全体においてもよく保存されており、近年の細胞周期及び癌研究の進展には酵母から得られた知見が大きな役割を果たしてきた。我々の研究室では真核細胞の細胞周期の研究に特異的阻害剤を利用し、その作用機構の解明に酵母遺伝学的手法を採り入れてきた。本研究では、さらに細胞周期阻害剤の探索にも酵母を利用し、得られた阻害剤を用いて真核生物の細胞周期に普遍的な制御機構を明らかにすることを目的として行った。 1.cdc変異株特異的阻害剤の探索 分裂酵母の細胞周期変異株としてcdc変異株が多く取得されている。cdc変異株は温度感受性変異株であり、非許容温度においては変異蛋白質の機能の失活により細胞周期の特異的な時期で生育が停止する。cdc変異株は許容温度では生育可能であるが、その変異遺伝子産物の機能は野生株に比べ低下しており、変異蛋白質自身、あるいは周辺の制御を標的とする薬剤に対して感受性が変化していることが予想される。本研究では細胞周期の中でも特にS期及びM期の開始の制御に重点を置き、G1/S期、G2/M期のcdc変異株の生育を特異的に阻害する物質の探索をペーパーディスクアッセイにて行った。その結果、Aspergillus属と思われるカビの培養液中にcdc25,cdc13など細胞分裂の開始に必須なCdc2キナーゼの活性制御に関わる変異株の生育を野生株に比べ極めて低濃度で阻害する活性を見出した。培養液をアセトンで抽出し、酢酸エチル抽出、シリカゲルカラム、ODS薄相クロマトグラフィーを行い、活性物質として抗真菌物質として報告されていたpreussinを同定した。 またcdc2株に対し著しい細胞伸長を引き起こすとともにcdc6,cdc7,cdc14など隔壁の形成に欠損を持つ株の生育を特異的に阻害する活性を未同定のカビの培養液中に見出した。精製の結果、この活性物質は各種癌細胞の形態を正常化する物質として本研究室のKwonらにより報告されたradicicolであった。 2.preussinの生物活性の検討(1)preussinの分裂酵母、培養動物細胞に対する生育阻害効果 preussinは分裂酵母野生株であるJY972株に比べ最も感受性の高かったcdc25株の生育を約1/500(最少阻止濃度MIC=10ng/ml)という低濃度で阻害した。この時細胞は異常に伸長した形態を示した。preussinは野生株に対しても最少生育阻止濃度(5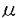 g/ml)において形態の伸長を引き起こした。ここで用いたcdc25株はcdc25遺伝子の温度感受性変異以外に栄養要求性変異としてura4,leu1,ade6などの変異を持つ。そこでこの感受性の差がcdc25遺伝子の変異に由来するものかを調べるため、cdc25単独変異株とura4,leu1,ade6の3つの栄養要求性を持つJY746株を掛け合わせた後、四分子解析を行った。その結果ura4,leu1,cdc25のそれぞれの単独の変異株は全て野生株に比べ感受性が上昇しており、もとのcdc25株の超感受性はこれらが合わさったものであると考えられる。それぞれの変異株のMICはcdc25で0.5g/ml,leu1で1g/ml、ura4で1g/ml程度であった。 g/ml)において形態の伸長を引き起こした。ここで用いたcdc25株はcdc25遺伝子の温度感受性変異以外に栄養要求性変異としてura4,leu1,ade6などの変異を持つ。そこでこの感受性の差がcdc25遺伝子の変異に由来するものかを調べるため、cdc25単独変異株とura4,leu1,ade6の3つの栄養要求性を持つJY746株を掛け合わせた後、四分子解析を行った。その結果ura4,leu1,cdc25のそれぞれの単独の変異株は全て野生株に比べ感受性が上昇しており、もとのcdc25株の超感受性はこれらが合わさったものであると考えられる。それぞれの変異株のMICはcdc25で0.5g/ml,leu1で1g/ml、ura4で1g/ml程度であった。 preussinは各種癌細胞に対し5-10g/mlのオーダーで増殖阻害を引き起こした。またラット正常線維芽細胞3Y1に対しても約10g/mlで細胞伸長とG1期での増殖の停止を引き起こした。 (2)preussinの細胞周期制御因子及びその他の酵素活性の阻害活性の検討 preussinはcdc25,cdc13などの細胞周期制御因子変異株の選択的な阻害物質として単離されたことからこれらの活性を直接阻害している可能性が考えられた。そこで分裂酵母野生株からp13suc1ビーズを用いて精製したCdc2キナーゼ、大腸菌で発現させ精製したヒトWee1キナーゼ及びGST-Cdc25Aを用いてそれらの酵素活性に対する阻害の有無を調べたところ、いずれの酵素に対してもpreussinは野生株のMICの10倍以上の高濃度においても阻害活性を示さなかった。またPKA,PKC,PTK,CAMK,EGFRなどの増殖に関わるプロテインキナーゼやトポイソメラーゼI,IIの活性もほとんど阻害しなかった。 またpreussinはロイシン要求性変異株の生育も強く阻害することから、ロイシンの取り込み及び蛋白質の合成に対する阻害効果を検討した。分裂酵母ロイシン要求性変異株に対し、[3H]ロイシンを取り込ませ細胞内及び蛋白質に取り込まれた放射線量を測定したがpreussinは阻害活性を示さなかった。 3.分裂酵母の遺伝学的手法を用いたpreussin標的分子の解明(1)酵母にpreussin耐性を付与する遺伝子の取得 先に述べたようにpreussinの阻害効果から予想される幾つかの酵素活性、生物活性について阻害の有無を調べてきたが、特定の阻害活性は見いだせなかった。そこで酵母の遺伝学的手法を用いてpreussinの標的分子の同定を試みた。preussinに最も感受性の高かったcdc25-3D株を宿主にしてLEU2遺伝子を持つプラスミドpDB248’をもとに構築した分裂酵母野生株のゲノムライブラリーから、宿主が生育できない0.1g/mlの培地にて選択を行い、いくつかの耐性クローン(pPSR1,pPSR2,pPSR4,pPSR6,pPSR7,pPSR9)を単離した。これらのクローンは調べた限り他の薬剤に対し耐性を付与せず、preussin特異的であった。 (2)pPSR1の解析 取得されたクローンの内、最も強い約100倍という耐性を付与したプラスミドpPSR1(PreuSsin Resistance)は同時にcdc25株の温度感受性も相補した。pPSR1の挿入DNA配列約10.5kbを解析したところ野生型のcdc25+遺伝子であり、preussin耐性を付与したのはこの遺伝子自身であると結論された。 (3)pPSR2の解析 プラスミドの制限酵素地図の比較から、重複した領域を含んでいたpPSR2とpPSR6の解析を行った。pPSR2は親株に対し約2-4倍の耐性を与えた。pPSR2は約4.5kbの挿入断片を含んでおり、塩基配列を決定したところ、約1.8kbのORFが見出された。またこのORFに含まれる形で裏側にも約1.4kbのORFが存在した。この表側1.8kbを含む約2.1kbをサブクローニングしたところpreussin耐性付与活性が見られた。このORFは出芽酵母の細胞質タイプの熱ショック蛋白質70ファミリーSSB1、SSB2と約70%と高い相同性を持ち、以前に本研究室の臼井によりK-252a耐性を付与する遺伝子として同定されたsks2+、及び神戸大学の久野らによりSSB1ホモログとして単離されたhsc1+と同一であった。以後この遺伝子をhsc1+と呼ぶことにした。 (4)その他のクローンの解析 pPSR4、pPSR9はpreussinに対し約2-4倍の比較的弱い耐性を与えるものである。pPSR4は約5.7kbの挿入断片を含んでおり、塩基配列を決定したところ出芽酵母のジスルフィドイソメラーゼに弱い相同性を持つORFと、タイプ2型プロテインホスファターゼと保存領域において相同性を持つORFの2つのORFが存在した。pPSR9は約6.9kbの挿入断片を含んでおり出芽酵母アラントインパーミアーゼと弱い相同性を持つORFが見出された。これらのクローンについては耐性付与に関わる領域の決定は行っていない。 4.hsc1+遺伝子の機能及びpreussin耐性付与機構の解析(1)hsc1+遺伝子の破壊 hsc1+遺伝子の機能を解析する目的で遺伝子破壊を行った。先に述べたようにhsc1+の読み枠中にはその裏側にも約1.4kbのORFが存在していたため、hsc1+遺伝子のN末領域の裏側ORFを含まない部位にPCRを用いて点変異を導入しBglII制限酵素の切断部位を作り、そこにura4+遺伝子断片約1.7kbを挿入し、hsc1+だけを破壊したコンストラクトを構築した。この断片を用いて二倍体野生株の形質転換を行った後、ura+となった株を選択し、サザンハイブリダイゼーションにより破壊を確認した。続いて四分子解析を行った結果、hsc1+は生育に必須な遺伝子ではなかったが、hsc1+破壊株は野生株に比べ若干生育が遅く、細胞の伸長が見られたことから細胞の増殖に何らかの異常があると考えられた。しかし温度感受性や低温感受性、金属イオンや浸透圧に対する感受性などは見られず、またpreussin感受性にも特に変化は見られなかった。次にcdc25温度感受性株とhsc1+破壊株を掛け合わせ、四分子解析を行ったところcdc25hsc1 ura4+の二重変異株は合成致死ではなかったが、液体培養において著しい細胞凝集と細胞の伸長や複数の隔壁が形成されるなどの生育異常が見られ、遺伝学的な相互作用が見られた。この形態はcdc25株に対し最少生育阻止濃度以上のpreussinを処理したものと類似しており、preussinの標的分子がhsc1+遺伝子産物自身である可能性が考えられる。Hsc1は熱ショック蛋白質70のメンバーであるが、熱ショックによって誘導されず、常温においても発現しており、通常の生育においても重要な機能を持つことが示唆される。Hsc1がCdc25と直接相互作用し、活性を制御している可能性が考えられるが、two-hybrid systemを用いた解析では両蛋白質間に直接の相互作用は検出されなかった。 ura4+の二重変異株は合成致死ではなかったが、液体培養において著しい細胞凝集と細胞の伸長や複数の隔壁が形成されるなどの生育異常が見られ、遺伝学的な相互作用が見られた。この形態はcdc25株に対し最少生育阻止濃度以上のpreussinを処理したものと類似しており、preussinの標的分子がhsc1+遺伝子産物自身である可能性が考えられる。Hsc1は熱ショック蛋白質70のメンバーであるが、熱ショックによって誘導されず、常温においても発現しており、通常の生育においても重要な機能を持つことが示唆される。Hsc1がCdc25と直接相互作用し、活性を制御している可能性が考えられるが、two-hybrid systemを用いた解析では両蛋白質間に直接の相互作用は検出されなかった。 5.preussin及びradicicol結合蛋白質同定の試み 分裂酵母感受性株にpreussin耐性を付与する遺伝子としてhsc1+を単離したが、hsc1+破壊株はpreussin感受性に特に変化がなかったことからHsc1によるpreussin耐性機構の詳細は不明である。そこでpreussinの一次標的分子を直接に同定するために、preussinのビオチン化誘導体をストレプトアビジンに結合させたアフィニティーカラムを用いてpreussin結合蛋白質の精製を試みた。preussinのビオチン化は20mgのpreussinから始め、ベンゼン環のニトロ化、還元によりp-位、o-位にアミノ基を導入した後、6-[(biotinoyl)amino]hexanoic acid,succinimidyl ester(biotin-SE)と縮合させることによりビオチン化preussin(p-を2.1mg,o-を1mg)を合成した(下図)。本合成品は活性の減少はみられたものの、元のpreussinと同様な生物活性を示した。現在このプローブを用いてヒト癌細胞であるHeLa細胞抽出液からpreussin結合蛋白質の精製を行っている。  6.まとめ 細胞周期、特に細胞分裂の制御の阻害剤の取得を目的として、分裂酵母cdc変異株特異的阻害剤の探索を行い、その結果preussin、radicicolなどの活性物質を同定した。preussinは分裂の制御因子であるcdc25変異株の生育を選択的に阻害する物質として同定したが、動物細胞に対してはG1期停止を引き起こすこと、またロイシンなどの栄養要求マーカーを持つ株も阻害することからpreussinの標的分子の機能に興味が持たれる。 preussin感受性株に耐性を付与する遺伝子として取得したhsc1+はそれ自身は生育に必須ではなく、破壊株もpreussin感受性にはあまり影響しない。しかしhsc1+とcdc25の間に遺伝学的な相互作用が見られたこと、またその時の形質がcdc25株にpreussinを処理したものと一部類似していたことから、preussinはCdc25とHsc1の相互作用を阻害することにより活性をあらわすというモデルが考えられる。しかし現在までCdc25とHsc1あるいはHsc1とpreussinの直接の相互作用は確認できておらず、三者の関係は不明である。preussin結合蛋白質の精製を目的として合成したビオチン化preussinはもとのpreussinに比べ約1/10〜1/20に低下していたものの、活性は保持しているため今後これを用いることにより標的分子の精製と同定が可能になったと考えられる。同様のことは、ビオチン化radicicolについてもいえる。 |
| 審査要旨 | | 酵母は単細胞で増殖する最も単純な真核生物であり、高等真核生物のモデルとして近年の細胞周期及び癌研究の進展に大きな役割を果たしてきた。本論文は、細胞周期阻害剤の探索に酵母を利用し、得られた阻害剤を用いて真核生物の細胞周期に普遍的な制御機構を明らかにした研究をまとめたものである。 1)cdc変異株特異的阻害剤の探索 分裂酵母cdc変異株の内、S期及びM期の開始の制御に関わる変異株の生育を特異的に阻害する物質の探索をペーパーディスクアッセイにより行い、一カビの培養液中にcdc25,cdc13など細胞分裂の開始に必須なCdc2キナーゼの活性制御の変異株の生育を選択的に阻害する物質としてpreussinを同定した。 2)preussinの生物活性の検討 preussinは最も感受性の高かったcdc25株の生育を野生株の約1/500(MIC=10ng/ml)という低濃度で阻害したが、ここで用いたcdc25株(cdc25-3 )はcdc25の変異以外にura4,leu1などの栄養要求性変異を持つことから、四分子解析により各変異を単独で、あるいは様々な組み合わせで持つ株を選択し、そのpreussin感受性を調べた。その結果ura4,leu1,cdc25のそれぞれの単独変異株は全て野生株に比べ感受性が高く、もとのcdc25株の超感受性はこれらが合わさったものであることが明らかになった。このうちleu1変異はpreussin感受性に最も影響が大きかったが、これはpreussinがロイシンの生合成ではなく細胞内への取り込みを阻害することによることが示された。 )はcdc25の変異以外にura4,leu1などの栄養要求性変異を持つことから、四分子解析により各変異を単独で、あるいは様々な組み合わせで持つ株を選択し、そのpreussin感受性を調べた。その結果ura4,leu1,cdc25のそれぞれの単独変異株は全て野生株に比べ感受性が高く、もとのcdc25株の超感受性はこれらが合わさったものであることが明らかになった。このうちleu1変異はpreussin感受性に最も影響が大きかったが、これはpreussinがロイシンの生合成ではなく細胞内への取り込みを阻害することによることが示された。 preussinはラット線維芽細胞3Y1に対しても約10g/mlで細胞伸長とG1期での増殖の停止を引き起こした。 preussinはその選択性から、Cdc2キナーゼ、Cdc25ホスファターゼ、Wee1キナーゼなどの活性を直接阻害する可能性が考えられた。そこで精製酵素を用いて試験管内での阻害実験を行ったところ、preussinはいずれに対しても阻害効果を示さなかった。 3)分裂酵母の遺伝学的手法によるpreussin標的分子の解祈 cdc25-3株を宿主に分裂酵母野生株のゲノムライブラリーから、preussin特異的に耐性を付与するプラスミドを単離した。この内100倍という最も強い耐性を付与したpPSR1は親株の温度感受性も相補し、塩基配列の解析の結果耐性遺伝子は野生型cdc25+遺伝子自身であった。 次に解析を行ったpPSR2は塩基配列の解析の結果、耐性遺伝子として出芽酵母の細胞質タイプのHsp70であるSSB1、SSB2のホモログとして報告されていたHsc1と同一であったため、以後この遺伝子をhsc1+と呼ぶことにした。 pPSR9はアラントイン酸パーミアーゼのホモログをコードするORFを含んでおり、psr9+と呼ぶことにした。 4)hsc1の機能とpreussin耐性付与機構の解析 hsc1+は遺伝子破壊の結果生育に必須ではなかったが、野生株に比べ若干生育が遅く細胞の伸長が見られた。cdc25 hsc1ura4+の二重変異株では細胞極性や隔壁形成の異常などそれぞれの単独変異では見られない形質を示した。この形態はcdc25株にpreussinを処理したものと類似していること、またこれらの形質はpreussin処理により強調されたことからcdc25とhsc1+の間に遺伝学的な相互作用が示されるとともに、この相互作用にpreussinが干渉する可能性が考えられた。cdc25株をG2期停止からのリリース後、分裂の開始を隔壁形成率を指標に調べたところ、preussin処理あるいはhsc1破壊により隔壁形成開始が遅延したことから、変異Cdc25の活性が回復する過程にHsc1が関与し、これをpreussinが阻害するというモデルが考えられた。 5)preussin結合蛋白質同定の試み preussin結合蛋白の同定を目指しpreussinのベンゼン環にビオチンを結合した誘導体を合成した。本合成品は活性は低下していたが、元のpreussinと同様な生物活性を示した。これを用いてHeLa細胞からpreussin結合蛋白質の精製を行い、約80kD及び50kd付近にビオチン化preussinを加えたときに特異的に現れるバンドを検出した。 以上、本論文は新しい細胞周期阻害剤を単離し、それを研究の道具として用いることによって細胞周期制御に新しい知見を加えるものである。これらの結果は、学術上貢献するところが少なくない。よって審査員一同は、本論文が博士(農学)の学位論文として価値のあるものと認めた。 |