細胞における生命反応はそれぞれのタンパク質を正しい場所に局在化してはじめて成り立つ。タンパク質の局在化は生化学・分子生物学・細胞生物学などの学問領域において最も重要な課題の一つとして研究が行われている。本論文は大腸菌の外膜リポタンパク質が内膜から遊離する分子機構を明らかにすることを目的として行われたものであり、序論と二章より成る。 序論では、リポタンパク質の局在化シグナル、これを認識するペリプラズムの分子シャペロン(LolA)、ならびに外膜に存在するリポタンパク質の受容体(LolB)の役割と相互作用についてこれまでの研究経過が述べられている。 第1章では、誤って内膜に局在化した主要外膜リポタンパク質(Lpp)の致死性が、ペプチドグリカンとの共有結合によって引き起こされることを明らかにしている。LppはN末端2番目(+2位)がSer(S)で、外膜に局在化する。C末端にはペプチドグリカンと共有結合するLys(K)をもつ。+2位のSerを、内膜局在化シグナルと考えられているAsp(D)に置換した変異体LppDKと、C末端のLysを欠失させ、Arg(R)がC末端となった変異体LppSRを作製した。さらに+2位とC末端の両方に変異をもつLppDRを作製した。野生型(LppSK)、LppSR、LppDRの発現は大腸菌の生育に影響をおよぼさなかったが、LppDKを発現させると速やかに生菌数が減少し、致死的影響をもたらした。LppDKを長時間発現させると、ショ糖密度勾配遠心法による内膜と外膜の分離ができなくなった。短時間の発現で調べた結果、LppDKとLppDRはどちらも内膜に局在化しており、+2位のアミノ酸が確かにLppの膜局在化シグナルとして働いていることが示された。さらに、外膜に局在化した野生型(LppSK)のみならず、内膜に局在化したLppDKもペプチドグリカンと共有結合していることを明らかにした。グロボマイシンによって外膜への移行を阻害すると、C末端にLysをもつLpp(LppSK)を発現させた時のみ致死的影響をもたらし、LppSRを発現する細胞は非感受性であった。以上のことから、Lppが内膜に蓄積すること自身は致死的ではないが、C末端にLysをもったLppが誤って内膜に局在すると、ペプチドグリカンとの間で共有結合がおき、結果的に細胞表層の崩壊をもたらすことによって致死性を示すと考察している。 第2章では、外膜リポタンパク質のキャリアータンパク質(LolA)に依存した内膜遊離機構の解析を行っている。リポタンパク質内膜遊離反応を単独で解析するために、点変異を導入した7種の変異体Lppを作製した。その中で、10番目のLeuをProに置換したLppL10Pを用いることによって、内膜遊離反応をright-side out膜小胞で解析できるin vitro実験系を構築した。内膜にLppL10Pを蓄積したスフェロプラストから、ATP存在下に調製したright-side out膜小胞にLolAを添加すると、LppL10Pが効率よく遊離した。ATPの非加水分解アナログや呼吸基質のみを用いた場合にはLppL10Pの遊離はみられなかった。さらに、外膜リポタンパク質の内膜からの遊離を担う因子の同定を進めるため、プロテオリポソームを用いた内膜遊離反応の再構成を行った。LolAならびにATPに依存した内膜遊離反応の再構成は、内膜タンパク質だけでなく、細胞質因子が必要であった。再構成実験系を用いて、内膜遊離反応に働く細胞質因子の精製を行った。部分精製した細胞質因子のN末端5アミノ酸の配列は無機ピロホスファターゼ(PPase)のものと一致した。さらに、PPaseの変異株あるいは酵母由来のPPase標品を用いた解析から、リポタンパク質の内膜遊離反応に関与する細胞質因子はPPaseであると結論した。一方、PPaseにATP分解活性はなかったので、ATP要求性は内膜因子によるものであると考えた。内膜遊離反応はP型ATPaseの阻害剤として知られるバナジン酸によって阻害されたので、内膜因子がP型ATPaseである可能性が考えられた。これらをもとに、ATPのエネルギーを駆動力とし、内膜因子とPPaseが協同して、外膜リポタンパク質の内膜遊離を行っていると考察している。 以上要するに本論文は、大腸菌のリポタンパク質が選別され、内膜あるいは外膜に局在化する機構について、細胞の構造維持に果たす役割ならびに内膜遊離反応に関与する新因子とエネルギー要求性について明らかにしたものであり、学術上・応用上寄与するところが少なくない。よって、審査員一同は本論文が博士(農学)の学位論文として価値あるものと認めた。 | 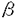 -ラクタマーゼとリポタンパク質の融合タンパク質を用いて、脂質修飾を受けるCysの隣のアミノ酸残基(+2位)がリポタンパク質の膜局在化シグナルとして働いており、ここがAspであるものは内膜に、それ以外のものは外膜に局在化することを示した。本研究の第1章では野生型を含めた4種の主要外膜リポタンパク質(Lpp)の変異体を用い、Lppの局在化シグナルを同定し、内膜に誤って局在化したLppがなぜ致死性を引き起こすかを解明した。
-ラクタマーゼとリポタンパク質の融合タンパク質を用いて、脂質修飾を受けるCysの隣のアミノ酸残基(+2位)がリポタンパク質の膜局在化シグナルとして働いており、ここがAspであるものは内膜に、それ以外のものは外膜に局在化することを示した。本研究の第1章では野生型を含めた4種の主要外膜リポタンパク質(Lpp)の変異体を用い、Lppの局在化シグナルを同定し、内膜に誤って局在化したLppがなぜ致死性を引き起こすかを解明した。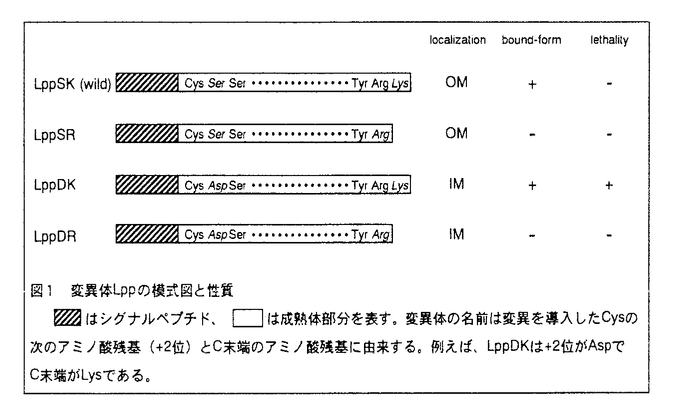
 はシグナルペプチド、
はシグナルペプチド、  は成熟体部分を表す。変異体の名前は変異を導入したCysの次のアミノ酸残基(+2位)とC末端のアミノ酸残基に由来する。例えば、LppDKは+2位がASpでC末端がLySである。
は成熟体部分を表す。変異体の名前は変異を導入したCysの次のアミノ酸残基(+2位)とC末端のアミノ酸残基に由来する。例えば、LppDKは+2位がASpでC末端がLySである。