胎盤は、代謝・免疫・内分泌等の多岐にわたる機能を有し、これらの機能は時期特異的に調節されている。本研究論文は胎盤機能の時期特異的な調節機構を調べることを目的とし、ラット胎盤の細胞膜タンパク質に着目し、主に二次元電気泳動法とタンパク質のアミノ酸配列を決定する手法を用いて解析を行っている。 約800種の膜タンパク質について二次元電気泳動にて検出し、これらより、妊娠の進行に伴い発現量の変化する150種の膜タンパク質を選びだした。これら150種には、ハウスキーピング遺伝子産物等も含むと考えられ、胎盤の時期特異的な機能に関わる分子の探索を行う必要性が考えられた。そこで、ホルモン処置により卵巣に新たに黄体を導入し、妊娠期間を延長させたラット(NCラット)では、妊娠後期にも関わらず、正常妊娠ラットの妊娠中期特異的な機能を回復することを利用し、正常妊娠ラットとNCラット妊娠20日目を比較し、時期特異的な機能に同期して発現する膜タンパク質の検出を試み、妊娠中期の機能に関わるpsM-I(MW76.4k,pI5.8)、psM-II(68.4k,5.5)、psM-III(44.2k,5.6)、および、妊娠後期特異的な機能に関わるpsL-I(MW36.2k,pI5.3)、psL-II(35.9k,5.3)、psL-III(27.0k,5.1)を検出した。 まず、妊娠中期特異的なpsM-IIIの解析を行った。psM-IIIは、妊娠18日目までほぼ一定のレベルで発現されており、妊娠20日目に検出限界以下となる。ラット胎盤由来の培養細胞であるRcho-1細胞では未分化な状態では発現しているが、分化すると消失し、細胞の増殖と分化の調節にも関与していると考えられた。Rcho-1未分化細胞を用い二次元電気泳動法でpsM-IIIを分離精製し、リジン残基特異的な酵素Achromobacterlyticus protease I(API)を用いて切断した後、得られた断片化ペプチドを逆相HPLC(C8カラム)で分離した。2種の明瞭なピークをアミノ酸配列分析に処した。得られたアミノ酸配列を基にデーターベース検索を行った結果、4種の高い相同性を有する分子(TDD5,Ndr1,Drg1,RTP)が検出された。これらは、いずれも近年報告された未知機能分子である。胎盤の時期特異的な機能調節に関わる未知な機能の分子の同定に成功した。 次に、psL-IとpsL-IIの解析を行った。二次元電気泳動像において、psL-Iに対しpsC-I、またpsL-IIに対しpsC-IIと、アイソフォーム様の分子の存在に気付き、これらを含め解析した。psC-I、-IIは、妊娠期全般にわたり発現しており、psL-I、-IIは妊娠16日目以降に発現していた。二次元電気泳動で分離精製した4種のタンパク質をAPIで処理した後、逆相HPLC(C18カラム)で分離した。ペプチドマップの結果、psL-IとpsC-I、およびpsL-IIとpsC-IIは、同一のタンパク質部位を有し、また、psL-IとpsL-IIおよびpsC-IとpsC-IIは、類似した一次構造を有する分子であることがわかった。これに加え、アミノ酸配列分析、質量分析の結果から、psL-IとpsC-Iは、Gタンパク質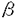 1サブユニット、またpsL-IIとpsC-IIは2サブユニットであることがわかった。これから、胎盤では、1,2サブユニットに翻訳後修飾がおきていることが考えられた。 1サブユニット、またpsL-IIとpsC-IIは2サブユニットであることがわかった。これから、胎盤では、1,2サブユニットに翻訳後修飾がおきていることが考えられた。 サブユニットの翻訳後修飾としては唯一、ヒスチジン残基がリン酸化されることが報告されているが、追試を行ったところ同修飾ではないことが解った。胎盤細胞で見られたサブユニットの修飾は、新規のものである。サブユニットの分子修飾は、胎盤以外の臓器(脳、卵巣、肝臓、腎臓)で調べたところ、脳でも見られた。従って、同修飾は上記以外の臓器でもおきている可能性が考えられた。Gタンパク質、 サブユニットは、近年、会合する多様な分子群が知られ、細胞内機能調節において重要な役割を果たしていることが明らかとなってきた。サブユニットのアミノ酸配列は、ヒト、ウシ、ラット、マウスで100%保存されており、種を超え同分子の高次構造維持が重要であると推察される。 サブユニットは、近年、会合する多様な分子群が知られ、細胞内機能調節において重要な役割を果たしていることが明らかとなってきた。サブユニットのアミノ酸配列は、ヒト、ウシ、ラット、マウスで100%保存されており、種を超え同分子の高次構造維持が重要であると推察される。 以上、本論文では、三量体Gタンパク質のサブユニットの分子修飾が妊娠後期胎盤に発現していること、妊娠中期に一連の特異遺伝子の発現のあることが明らかにされ、胎盤の機能調節に関する新たな知見が加わった。特にGタンパク質の分子修飾については他の組織での重要性も推定され重要な発見である。よって、審査員一同は本論文が博士(獣医学)の学位論文として価値あるものと認めた。 |  サブユニットの分子修飾
サブユニットの分子修飾 、
、