イノシトールリン脂質(PI)情報伝達系は、ホルモンの分泌、神経伝達物質の放出、細胞増殖因子による増殖など重要な生命現象に関わる重要な情報伝達経路である。ジアシルグリセロールキナーゼ(DGK)は、PI情報伝達系の酵素の1つで、ジアシルグリセロール(DG)をリン酸化し、フォスファチジン酸(PA)を産生する。DGとPAは共に脂質性セカンドメッセンジャーとして機能している。DGはプロテインキナーゼC(PKC)の活性化を、PAはrasGTPase活性化タンパク質やPI-4-キナーゼの活性調節を行っている。さらにDGとPAはmitogenとしても機能している。したがって、DGKはこれらセカンドメッセンジャーの細胞内濃度を調節するという役割を担う重要な酵素と考えられている。しかし、生理機能がまだ十分に解明されておらず、今後の課題となっている。最近、相次いで哺乳類からDGKアイソザイムが単離された。現在、アイソザイムごとの組織発現の同定、活性化因子や下流因子の同定、さらにどのような細胞機能に関わっているか解明が待たれている。 脊椎動物の視細胞の1つの桿体では、光受容機構にcGMPカスケードが使用されていることは明らかにされているが、錐体の光受容機構は明らかにされていない。一方、ショウジョウバエでは、視覚系突然変異体の詳細な解析から、PI情報伝達系が光受容に関与していることが示されている。生化学的解析から、脊椎動物網膜において光に依存したPI代謝の活性が検出され、さらに近年、ショウジョウバエ視細胞特異的分子のホモローグが、脊椎動物の視細胞で発現していることが報告された。これらのことから、脊椎動物視細胞でもPI代謝系が機能している可能性が示唆された。 これまでに我々の研究室において、ショウジョウバエの視覚系突然変異体、retinal degeneration A(rdgA)の解析を行い、視細胞特異的DGK2(rdgA遺伝子産物)が視細胞の光受容膜に局在し、光受容、及び、視細胞の構造維持に必要なことを明らかにした。本研究はこのrdgA遺伝子を手掛かりとして、ラット網膜で発現しているDGK遺伝子(DGK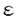 )を単離し、その発現パターンをin situハイブリダイゼーション法と抗体染色法によって明らかにしたものである。 )を単離し、その発現パターンをin situハイブリダイゼーション法と抗体染色法によって明らかにしたものである。 はじめに、ラット網膜で発現しているDGKの単離を試みた。RT/PCR解析の結果から、新規のDGK(DGK)が発現していることが示されたので、ラットDGK遺伝子のcDNAの全長を単離し、塩基配列と予想されるアミノ酸配列を決定した。この遺伝子は、全長約3.3kb、567アミノ酸をコードし、DGKの分子量は約64kDと予想された。この酵素は、これまでに単離されているDGKすべてに保存されているzinc-finger構造と触媒領域を持つ新規のアイソザイムで、ヒト精巣から単離されたDGK(ヒトDGK)と高い相同性を示した。ヒトDGKはPI代謝によって産生されるDGを特異的な基質とすることから、ラットDGKも同様な基質特異性を示し、PI情報伝達系で機能していると予想された。DGK遺伝子は、その発現パターンから網膜や脳で重要な機能を担っていることが示唆されたので(後述)、DGK遺伝子のin vitroでの機能を知ることは重要であること考え、マウスからDGKのホモローグの単離を行った。マウスDGKcDNAは全長約3.3kbであり、ラットDGK同様、567アミノ酸、分子量は約64kDのDGKタンパク質をコードしていると予想された。 次にラットとマウスDGKの機能を解析した。ノーザンハイブリダイゼーションにより、DGKは網膜で非常に強く発現し、脳にも発現していることが分かった。これまでにラット脳から単離されている4つのDGK(DGKI〜IV)と網膜における発現量を較べてみたが、DGKmRNAのみが網膜での強い発現を示した。DGKmRNAの組織内分布を調べるために、in situハイブリダイゼーションを行った。網膜では、視細胞の細胞体が存在しているouter nuclear layer(ONL)と2次ニューロンである双極細胞などの細胞体が存在するinner nuclear layer(INL)でシグナルが検出された。脳では、小脳のプルキンエ細胞と顆粒細胞、海馬のAmmon’s hornの顆粒細胞やdenate gyrusの錐体細胞で発現していた。 DGKタンパク質の発現を調べるために、DGKに対する抗体を作製した。組織切片の抗体染色の結果、網膜ではDGKタンパク質は、視細胞外節に局在していることが明らかとなった。その他、outer plexiform layer(OPL)やinner plexiform layer (INL)でも発現していた。 本研究では、網膜で強く発現しているDGK遺伝子をラットとマウスから単離し、発現パターンを解析した。網膜では、光受容が行われる視細胞外節に多く局在し、光情報伝達への関与が示唆された。光情報伝達の停止機構にPKCが関与していることから、DGKタンパク質がDGの量を調節しPKC活性を制御している可能性が考えられる。また、ショウジョウバエrdgAタンパク質のように、PA産生の調節を行い、視細胞外節の構造維持のための脂質供給に関わっている可能性も考えられる。この場合、脊椎動物と無脊椎動物の視細胞で用いられている機構の共通性を示しており興味深い。脳でのDGKmRNA発現パターンは、mGluR1やPKC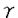 と類似しており、mGluR1はPI代謝系に連結していることから、DGKもこのカスケードの一員である可能性がある。mGluR1とPKCはシナプスの可塑性に関与していることから、DGKは脳でも重要な役割を担っていると予想される。 と類似しており、mGluR1はPI代謝系に連結していることから、DGKもこのカスケードの一員である可能性がある。mGluR1とPKCはシナプスの可塑性に関与していることから、DGKは脳でも重要な役割を担っていると予想される。 | 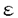 の単離と解析
の単離と解析