最近作られたエンドセリン(ET)-1遺伝子欠損マウスは正常マウスより高血圧を示し、ゆえに、内因性のET-1は血圧調節因子の一つであることが証明された。しかしながら、ET-1は血管内皮細胞由来の血管収縮因子として発見された分子であるので、この結果は予想とは逆のものであり、ET-1の欠損が如何にして高血圧をもたらすのかについては不明であった。そこで、ET-1の欠損が高血圧をもたらす機序を明らかにする目的で本研究を行った。本研究ではET-1欠損マウスの交感神経活動に特に注目した。その理由は、1)ET-1が中枢神経系内にも存在し、中枢性循環調節に関与することを示唆する証拠がすでに多く集まっていること、2)交感神経系は中枢性循環調節の主要な出力経路の一つであること、3)ET-1欠損マウスでは交感神経活動が亢進している可能性を示唆する間接的な証拠がすでに得られていたこと、である。具体的には、以下の2点を目標とした。 1)マウスの腎交感神経活動を直接測定する方法を確立し、マウスとラットにおいて、腎交感神経活動の基本性質と、その動脈圧受容器反射と化学受容器反射による調節を比較すること。 2)ET-1欠損マウスとその同腹兄弟の野生型マウスを用いて、同様に、腎交感神経活動とその調節を比較すること。 本研究で得られた主要な知見を以下に記す。 1.ウレタンで麻酔し、人工呼吸したマウスの腎交感神経活動を、世界で初めて記録することが出来た。 2.安静時の腎交感神経活動は、マウスの方がラットより有意に少なかった。腎神経に含まれる軸索数の差であると考えられた。フェニレフリンまたはニトロプルシッドの静脈内投与によって血圧を変動させた時の平均血圧-腎神経活動関係をシグモイド曲線に当てはめて動脈圧受容器-腎神経反射の特性を調べた。その結果、腎交感神経活動の最大値と動作範囲および平均血圧の中央値(MAP50)はマウスの方が有意に小さく、曲線の傾きはマウスの方が有意に大であった。 3.同様に、平均血圧-心拍数関係をシグモイド曲線に当てはめて動脈圧受容器-心拍数反射の特性を調べたところ、安静時心拍数と最大および最小心拍数はマウスの方が有意に大きく、曲線の傾きはマウスの方が有意に大であった。これらの動脈圧受容器反射の差異は種差に由来するものと考えられた。 4.人工換気中の二酸化炭素濃度の増加および酸素濃度の減少によってひき起こされた、化学受容器反射による腎交感神経活動の増大反応の大きさは、マウスとラットで差がなかった。 5.ET-1欠損マウスと野生型マウスを比較したところ、安静時において、血圧と腎交感神経活動が、ET-1欠損マウスの方が野生型マウスより有意に大であった。心拍数には差がなかった。 6.ET-1欠損マウスと野生型マウスの比較において、動脈圧受容器-心拍数反射の特性には差がなかった。しかしながら、動脈圧受容器-腎神経反射曲線は、安静時の血圧と腎交感神経活動の亢進および動作範囲の増大を反映して右上方にリセットされていた(図1)。 7.低酸素負荷による腎交感神経興奮反応はET-1欠損マウスと野生型マウスで差がなかった。いっぽう、二酸化炭素負荷による腎交感神経興奮反応はET-1欠損マウスの方が有意に減弱していた。 これらの結果から、内在性のET-1は、中枢神経による二酸化炭素の検出に関与していると考えられた。一方、私の属する研究グループの以前の報告によると、ET-1欠損マウスでは、無麻酔安静時の血中酸素分圧の低下と、低酸素負荷と二酸化炭素負荷による横隔神経を出力とする化学受容器-呼吸反射の双方の減弱が観察された。これらをあわせ考えると、ET-1欠損マウスにおける高血圧の原因は以下のように推測できる(図2)。 1)何らかの原因によって血中酸素分圧の低下が生じても、呼吸反射の異常により正常酸素分圧への是正が行われずに低酸素状態が維持されてしまう。 2)低酸素状態は、正常に保たれている末梢化学受容器-腎交感神経反射によって腎交感神経を興奮させ、もって血圧を上昇させる。 3)動脈圧受容器反射は存在はしているが、亢進した腎交感神経活動の動作域に順化(リセット)しているために、安静レベルの血圧と交感神経活動を正常動物レベルにまで押し下げる能力を失っている。 本研究により、ET-1欠損マウスにおける高血圧の機序に関し、合理的な説明が可能になった。また、マウスの腎交感神経活動を定量する方法の確立は、ET-1欠損マウスに限らず、応用の可能性の高いものである。  図1.エンドセリン-1欠損マウスにおける平均血圧(MAP)腎交感神経活動(RSNA)関係のリセッティング。丸印は安静時の平均値を、垂直線は最大時、安静時、最小時の各々における標準誤差を示す。*P<0.05。 図1.エンドセリン-1欠損マウスにおける平均血圧(MAP)腎交感神経活動(RSNA)関係のリセッティング。丸印は安静時の平均値を、垂直線は最大時、安静時、最小時の各々における標準誤差を示す。*P<0.05。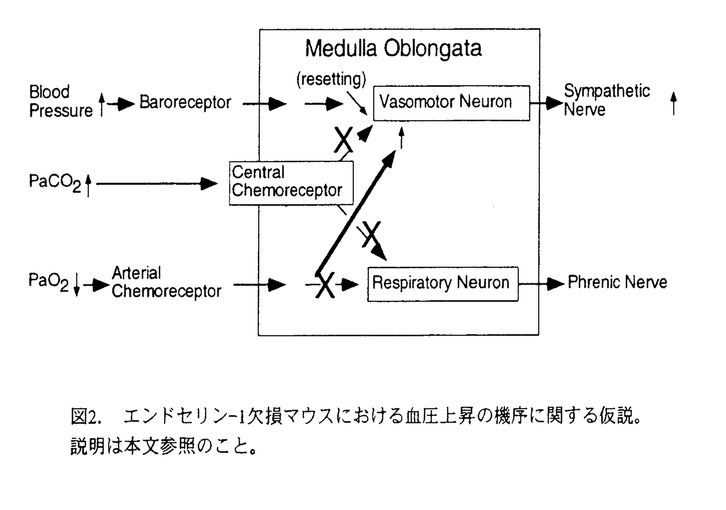 図2.エンドセリン-1欠損マウスにおける血圧上昇の機序に関する仮説。説明は本文参照のこと。 図2.エンドセリン-1欠損マウスにおける血圧上昇の機序に関する仮説。説明は本文参照のこと。 |