本研究は細胞内イノシトールリン脂質代謝で主役を演じるイノシトールリン脂質特異的ホスホリパーゼC(PLC)のアイソザイムの1つPLC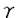 1の機能を明らかにすることを目的として行われたものである。著者は第783位のチロシンでリン酸化されたPLC1を認識する抗体(抗PLC1PY抗体)を作成し、それを用いて、細胞増殖時の形態変化やDNA合成に、PLC1の783位のチロシンリン酸化がどの様に関与するかを解析し、以下の結果を得ている。 1の機能を明らかにすることを目的として行われたものである。著者は第783位のチロシンでリン酸化されたPLC1を認識する抗体(抗PLC1PY抗体)を作成し、それを用いて、細胞増殖時の形態変化やDNA合成に、PLC1の783位のチロシンリン酸化がどの様に関与するかを解析し、以下の結果を得ている。 1、細胞を抗PLC1PY抗体とrhodamine phalloidinで二重染色したところ、PDGF刺激時には、PLC1の783位がチロシンリン酸化を受けると共に、リン酸化したPLC1は細胞骨格画分と細胞膜のラッフリング部位に局在する様になるのが観察された。細胞を抗PLC1抗体と抗PLC1PY抗体で二重染色した結果からは、PDGFの刺激によって第783位がチロシンリン酸化されたPLC1は細胞骨格に多く局在していることが明らかになった。このことから、PLC1の783位のチロシンリン酸化とPLC1の細胞骨格へのトランスロケーションが相関することが判明した。イムノブロットの結果からは、PDGF刺激前には、PLC1は細胞骨格と可溶化画分の両方に存在するが、PDGF刺激した後30分には可溶化画分に存在する多くのPLC1が細胞骨格画分へトランスロケーションし、90分後には、PLC1は刺激する前の状態に戻ること、又、PDGF刺激30分での細胞骨格画分に局在するPLC1の783位のチロシンの著しいリン酸化がみられ、90分経つと、リン酸化されたPLC1がかなり減っていることが観察された。これらの結果より、PLC1の783位のチロシンリン酸化がPLC1の細胞骨格へのトランスロケーションと相関していることが強く示唆された。 2、PDGF刺激した後、正常細胞では、アクチン線維の切断と細胞膜ラッフリングが見られるが、抗PLC1PY抗体を微量注入した場合は、PDGF刺激によるアクチン線維と細胞膜の変化は完全に阻害された。PLC1のSH2領域はPLC1へのシグナルに対しdominant negativeに働き、PLC1のSH2領域を細胞内に注入すると、抗PLC1PY抗体の微量注入と同じように、PDGFによるアクチン線維の切断と細胞膜ラッフリングを阻害した。これらの結果から、PLC1の783位のチロシンリン酸化は細胞骨格の再構築を調節していることが明らかになった。 3、細胞内にチロシン783リン酸化ペプチドを細胞内に微量注入し、細胞形態、細胞骨格系への影響を調べたところ、予想に反して、PDGF刺激と同じように、細胞内アクチン線維の切断と細胞膜ラッフリングが誘導された。更に、リン酸化ペプチドは細胞骨格と細胞膜ラッフリング部位に局在していることも判明した。これらの結果から、PLC1の783位のチロシンリン酸化が細胞骨格の再構築に重要な役割を担っていることが示された。 4、抗PLC1PY抗体を細胞内に微量注入したところ、PDGFによるDNAの合成がほぼ完全に抑制された、すなわち、PLC1は増殖因子PDGFの下流に存在し、PLC1の783位のチロシンリン酸化はDNAの合成にも重要であることが判明した。 以上、本論文は、PDGF刺激後、PLC1の783位のチロシンリン酸化され、リン酸化されたPLC1は細胞骨格と細胞膜ラフリング部位に局在すること、PDGFによるアクチン繊維の切断と細胞膜ラフリングは抗PLC1リン酸化抗体の微量注入よりブロックされ、DNA合成も抑制されること、783位のチロシンリン酸化含むPLC1ペプチドの微量注入はPDGF刺激と同じようにアクチン繊維の切断と細胞膜ラッフリングを引き起こすことを明らかにしている。本研究は、PDGFによるPLC1の783位のチロシンリン酸化は細胞増殖のみならず細胞骨格の再構築に関与していることを解明していく上で、重要な貢献をなすと考えられ、学位の授与に値するものと考えられる。 | 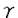 1の783位のチロシンリン酸化は細胞骨格の再構築を調節する
1の783位のチロシンリン酸化は細胞骨格の再構築を調節する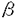 ,
,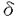 の三種類に分類されているが、その中でPLC
の三種類に分類されているが、その中でPLC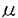 gの合成したリン酸化ペプチドGFpYVEANPMPTFK(PLC
gの合成したリン酸化ペプチドGFpYVEANPMPTFK(PLC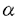 -アクチニン、ビンキュリンに結合したPIP2が減少することも判明している。PLC
-アクチニン、ビンキュリンに結合したPIP2が減少することも判明している。PLC