本研究はチロシンキナーゼの生物学的機能を明らかにするため、腫瘍細胞において癌遺伝子として同定された新規チロシンキナーゼalkの癌原遺伝子cDNA全長の単離、同定を試み、正常型ALKの構造の決定と、発現機能の解析を試みたものであり、下記の結果を得ている。 1. 2;5染色体転座を伴うヒトリンパ腫の一群において、転座の結果として生じるnpm-alk融合遺伝子産物は、既知の核タンパク質nucleophosmin(NPM)の一部とN末側を欠いた新規チロシンキナーゼALKとの融合タンパク質であるが、この新規チロシンキナーゼの正常型alk癌原遺伝子同定のため、npm-alk cDNA断片をプローブとして、マウス及びヒトalk cDNAの全長をクローニングした。マウスalk及びヒトalk遺伝子は、それぞれ1621,1619アミノ酸残基のタンパク質をコードし、予想されるマウスALKタンパク質は1014アミノ酸残基からなる細胞外領域、疎水性の27アミノ酸残基からなる膜貫通領域、そして保存された細胞内チロシンキナーゼドメインを含む560アミノ酸残基の細胞内領域により構成される典型的な受容体型チロシンキナーゼであった。ALKタンパク質のチロシンキナーゼドメインはインシュリンレセプターファミリー、とりわけLTK(leukocyte tyrosine kinase)と高い相同性を示し、ALKの細胞外領域においても、アミノ酸残基687番目から1034番目の膜貫通領域までが、LTKの細胞外領域と約50%(amino acid identity)の相同性があり、ALKとLTKが新たなサブファミリーを形成していることが分かった。LTKと相同性のある領域より更にN末端側の配列は既知のタンパク質の配列と類似せず、Ig-likeドメイン、ファイブロネクチンtypeIIIドメインなど、他の受容体の細胞外領域に多く見られるモチーフは検出されなかった。 2. ALKタンパク質に対する抗体(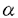 -ALK)はalk発現プラスミドを導入したCOS細胞のlysate中に約220kDaのタンパク質と特異的に反応した。また、-ALKによる免疫沈降物のin vitroキナーゼ反応を行い、ALKタンパク質のキナーゼ活性を検討したところ、alk発現プラスミドを一過性に導入したCOS細胞の溶解物中の-ALK免疫沈降物からは、分子量約220kDaのチロシンリン酸化タンパク質が検出された。このことは、alk遺伝子産物が自己リン酸化能があること示している。 -ALK)はalk発現プラスミドを導入したCOS細胞のlysate中に約220kDaのタンパク質と特異的に反応した。また、-ALKによる免疫沈降物のin vitroキナーゼ反応を行い、ALKタンパク質のキナーゼ活性を検討したところ、alk発現プラスミドを一過性に導入したCOS細胞の溶解物中の-ALK免疫沈降物からは、分子量約220kDaのチロシンリン酸化タンパク質が検出された。このことは、alk遺伝子産物が自己リン酸化能があること示している。 3. ノザンハイブリダイゼーション解析によるalk mRNAの各組織における発現の解析では、マウス新生仔脳(1日齢)において約9kbのalk mRNAが検出された。また、4週齢のマウス脳においても、新生仔脳より低い発現量で、同じく約9kbのalk遺伝子転写産物が検出された。脳以外の組織における発現は認められなかった。各週齢別のマウス脳におけるALKタンパク質の発現を、-ALKを用いたウエスタンブロットによって検討した結果では、ALKタンパク質は1週齢の脳において最も高い発現を示し、その後週齢を追って発現量の減少が見られた。ALKタンパク質の発現は、3週齢近辺で最小であり、その後少なくとも1年齢まで発現レベルに変化は認められなかった。マウスの脳においては、出生後10日までニューロンの伸張、シナプス連絡の形成が行われているが、こうした中枢神経系の発達段階、ニューロン網の形成、発達期において、ALKは重要な役割を果たしていることが考えられた。加えて、ALKは成熟した脳においても低いレベルで発現し続けていることから、ニューロンの生存、維持に関わっているとも予想された。 4. alk anti-sense cRNAをプローブとして用いたマウス胎仔(E15及びE19)及び一週齢マウス脳組織切片のin situハイブリダイゼーション解析では、胎仔において、中枢神経系、末梢神経系に限局してalkが検出された。脳においては、脳幹、中脳、間脳、及び終脳の細胞集団と一致して発現が見られ、脊髄ではその腹側、運動ニューロン群が存在する領域に高い発現が見られた。末梢神経系では、三叉神経節、上頸神経節(SCG)、後根神経節(DRG)、腸管軸に沿った自律神経系の末端神経節において発現が見られた。一週齢マウス脳におけるalk mRNAは限局した発現分布を示し、嗅球(僧帽細胞)、視床の一部神経核(背外側核、束傍核、不確帯)、中脳(赤核、上丘)のニューロンで強く検出された。生後一週間のマウス脳において、こうした終脳皮質と末梢器官を中継する神経核に限局してalkの発現が見られることは、シナプス形成が活発な時期のニューロンにおいて、ALKが機能していることを示唆する。中枢神経系におけるalkの発現解析では、少なくとも、ニューロンの分化とシナプス形成を伴う成熟した特定の神経核の形成期においてALKが何らかの役割を果たしていることが示唆された。 以上、本研究において新規に単離、同定された受容体型チロシンキナーゼalkの発現機能の解析により、神経系特異的に発現するこのチロシンキナーゼの機能の研究の端緒を開いた。本研究は受容体型チロシンキナーゼの関わる神経系の形成、発達、維持のメカニズム等の研究において重要な貢献をなすと考えられ、学位の授与に値するものと考えられる。 |