| 背景・目的 C型肝炎ウイルス(以下HCV)は、フラビウイルス科の独立した一属に分類される、一本鎖プラス鎖RNAウイルスである。HCVは輸血後非A非B肝炎の主要な病原体であり、感染者には慢性感染の後に高率に肝硬変・肝細胞癌が発生する。全長約9,500塩基のゲノムRNAは341塩基前後の5’非翻訳領域、3,008-3,037残基のポリペプチドをコードする翻訳領域、200-300塩基の3’非翻訳領域よりなり、3’非翻訳領域はポリU/C配列の後、98塩基の3’Xと呼ばれる構造で終わる。翻訳の開始は、RNAの5’部位に存在するinternal ribosomal entry site(IRES)に依存する点で通常の真核細胞mRNAに見られるcap依存性の蛋白合成と異なる。合成されたポリペプチドは宿主細胞のシグナルペプチダーゼ、ウイルスがコードする二種類のプロテアーゼによりプロセシングを受け、個々の蛋白となる。 IRESが最初に同定されたピコルナウイルスでは、リボゾームは5’非翻訳領域内の数百塩基の塩基配列を認識し、RNA上にエントリーする。HCVの5’非翻訳領域は開始コドンとならないAUGが複数(2-5個)存在し、複雑な二次構造を形成しうる点でピコルナウイルスと類似している。実際に1992年にTsukiyama-KoharaらによってHCV5’非翻訳領域内110塩基から341塩基にIRESが存在することが示され、その後シスエレメントの解析が多くのグループにより行われてきた。5’非翻訳領域内に欠失や置換を導入する実験により、5’端に形成が予想されるヘアピン構造を除き、ほとんどの領域が活性発現に必要であることが示唆されている。5’端のヘアピン領域については報告によってIRES活性に必要であるとするものと、阻害的に働くとするものがある。また、IRESの5’境界については、110塩基、40塩基前後、28塩基、1塩基とする報告があり、混乱が見られる。一方、3’境界についても、5’非翻訳領域を直接レポーターの翻訳領域に接続してIRES活性が見られる場合と、HCVコア蛋白の31塩基を含めないと十分な活性が得られない場合、さらにポリオウイルスキメラを用いた実験系でコア蛋白のN末2/3がウイルス複製を活性化する場合が報告されている。 生物活性が異なるウイルス株間での組換え実験は、機能解析に重要な役割を果たしてきた。HCVには6つ以上の遺伝子型が存在するが、この中で1b,2bの5’非翻訳領域は90%以上の相同性があるにもかかわらず、5’非翻訳領域の5’端から58塩基を欠くRNAを用いた無細胞蛋白合成系での比較では2bのIRES活性の方が高いことが観察されていた。すでにこの2つの株間の組換え実験が無細胞蛋白合成系で行われているが、IRESの5’側を欠いているため、真の結果ではない可能性があり、全長を用いた組換え体の活性を調べる必要がある。 本研究では、IRESの機能発現に必要な領域を特定するために5’非翻訳領域全域について遺伝子型1b,2b間の組換え実験を行った。IRESの範囲を特定することは、今後HCV IRESの解析を進めていく上で必要不可欠な実験であり、IRES機能発現に必要な構造を評価することにもつながる。遺伝子型1bのRNA5’部位約900塩基にわたり欠失変異体を作製し、IRESの5’境界と3’境界を決定した。 方法 IRES部位に様々な変異を含むジシストロニックRNAを作製し、無細胞蛋白合成系と培養細胞でIRES活性を測定した。 ジシストロニックRNAを用いることで、培養細胞へのトランスフェクションの効率を補正することが可能となる。第一シストロンにはインターナルマーカーとなるクロランフェニコールアセチルトランスフェラーゼ(CAT)、第二シストロンにはHCV RNAを含む組換え体を用いた(組換え体と5’境界の解析にはコア蛋白、3’境界の解析にはgreen fluorescence protein(GFP)と大腸菌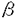 ガラクトシダーゼを用いた)。 ガラクトシダーゼを用いた)。 IRES領域については、最初に遺伝子型1b,2bの5’非翻訳領域全長を含むジシストロニックcDNAをオリゴデオキシリボヌクレオチドを用いて作製し、組換え体は5’非翻訳領域内の制限酵素部位を利用して作製した。5’欠失体・3’欠失体は、RNAの予測二次構造あるいはコア蛋白の機能ドメインを参考に、PCRを用いて遺伝子型1bについて作製した(別図参照)。 無細胞蛋白合成系としてはウサギ網状赤血球抽出液(RRL)、HeLa細胞抽出液(HeLa S10)を用い、35Sメチオニン存在下で合成された蛋白をSDS-PAGEで分離し、第二シストロンの産物をイメージアナライザーで定量した値をIRES活性とした。 培養細胞系としてHeLa,AGMK,HepG2細胞を用いた。各細胞へはリポフェクチンによるRNAトランスフェクションにより遺伝子を導入した。第一シストロンの産物はCAT活性として、第二シストロンはELISA法によりコア抗原量として定量し、コア抗原量をCAT活性で割った値を培養細胞中でのIRES活性とした。 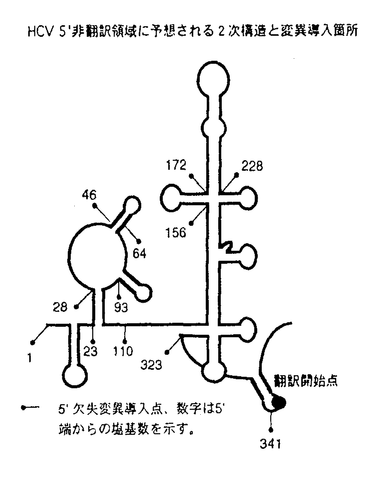 HCV5’非翻訳領域に予想される2次構造と変異導入箇所 HCV5’非翻訳領域に予想される2次構造と変異導入箇所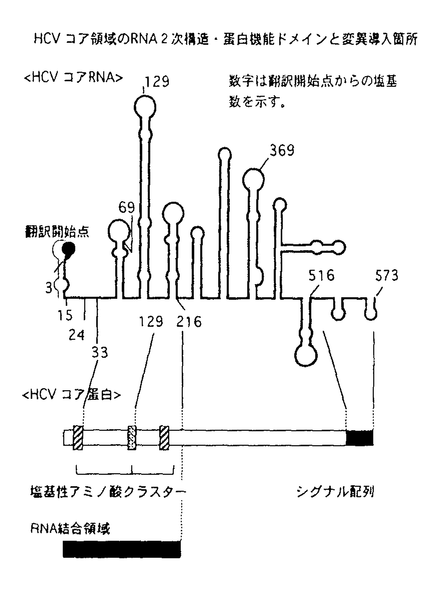 HCVコア領域のRNA2次構造・蛋白機能ドメインと変異導入箇所結果・考察 HCVコア領域のRNA2次構造・蛋白機能ドメインと変異導入箇所結果・考察 3種類の培養細胞で遺伝子型1bと2bのIRES活性を比較した。用いたすべての細胞で2b IRESの方が1b IRESよりも高い活性を示した。活性の差はHeLa,HepG2細胞では2倍、AGMK細胞では5倍であり、細胞によりIRES関連因子の分布が異なることが予想された。 5’非翻訳領域を3つの領域に分けて作製した1b,2b間の組換え体は、無細胞蛋白合成系では2bと同等かそれよりも高い活性を示し、培養細胞系では1b,2bの中間の活性を示した。組換え体の示すパターンからは、特定の領域が活性差を決定しているのではなく、3つの領域すべてが活性に関与し、さらに領域間に相互作用が存在することが考えられた。IRES機能の発現にはRNAの高次構造が重要であることが予想された。 遺伝子型1bを用いて、IRESの5’境界を決定した。無細胞蛋白合成系・培養細胞系ともに、5’端から27塩基の欠失により、無細胞蛋白合成系では1.4倍、培養細胞系では2倍以上IRES活性が上昇し、これ以上の欠失ではIRES活性は徐々に低下した。無細胞系では110塩基まで欠失したRNAでも弱い翻訳活性が認められたが、156塩基以上の欠失では活性は消失した。IRESを最大量のリボゾームをmRNAにエントリーさせる領域と定義すると、この結果はIRESの5’境界は28塩基から45塩基の間にあることを示している。一方、最低限エントリーさせる領域とすると、5’境界は無細胞系では110塩基から155塩基の間に存在することになる。5’端のヘアピン形成部位は無細胞蛋白合成系では抑制的に働くが、培養細胞系ではHeLa細胞では活性に変化を与えず、AGMK細胞ではむしろ欠失により活性が低下した。この結果は、この部位を含むRNA配列に作用する宿主因子が存在し、その分布が細胞により異なると考えると説明可能である。 3’境界を決定する際のウイルスRNA由来塩基配列後の核酸配列の効果を、GFPをレポーターとして5’非翻訳領域に直接つないだ場合と、30塩基のベクターの配列を開始コドンの直後に挿入した場合とで比較した。直接つないだ場合はIRES活性が検出できるのに対し、介在配列を挿入した場合は検出できなかった。しかし、後者に対してコアN末の塩基配列を24塩基挿入すると活性はほぼ同等となり、33,69塩基の挿入で各5.1,3.8倍に上昇した。以上よりコアの塩基配列33塩基が必要であると考えられ、また、レポーターの塩基配列もIRES活性に影響することが示唆された。 種々の長さのコア領域の3’欠失体を作製し、レポーターであるガラクトシダーゼmRNAの上流につなぎ、無細胞蛋白合成系で3’境界を検討した。33塩基までの挿入は翻訳活性を上昇させた。さらに69塩基までの挿入はRRLでは活性を上昇させたが、HeLa S10では抑制した。70塩基から129塩基の挿入で翻訳産物量は低下するが、130塩基から516塩基の挿入で上昇し、369,516塩基を挿入した場合現在3’境界と考えられている33塩基を挿入した場合よりも高い活性を示した。同じ融合蛋白を、HCVの5’非翻訳領域の大半を欠失させて5’端依存的に合成させた場合は、コア領域の挿入によるこのような活性の変化はみられないことから、HCV5’非翻訳領域と翻訳領域の間に相互作用があることが示唆された。これらの結果から、IRESの3’境界は370塩基から516塩基の間にあると考えられた。翻訳領域がIRES活性に影響を与える機序としては、RNAの構造として重要である可能性とコア蛋白自身が重要である可能性の両方が考えられる。 結論 遺伝子型2bのIRESは1bよりも高い活性を示した。組換え体の活性パターンは、IRES内の特定の領域ではなく全体の領域が活性に影響を及ぼすことを示し、RNAの高次構造が重要であることが示唆された。IRESの5’端は28塩基と45塩基の間に、3’境界は翻訳領域の370塩基と516塩基の間にあることが明らかとなった。5’端の欠失体の示すIRES活性や2b,1b間の活性比には細胞差があり、IRES関連因子とIRESの相互作用は細胞による差があることが考えられた。 |