大腸菌のP1ファージがコードしている部位特異的組換え酵素Creは標的配列である34bpのloxP配列を特異的に認識し、その標的配列間で相同組換えを効率よく行う。標的配列loxPは、両端各13塩基は相同でパリンドローム構造をとり、その構造の間に8塩基(ATGTATGC)の非対象なスペーサー配列がある。組換えの際に、このスペーサー内で中央部6塩基が、cohesive末端となるように切断され、切断されたDNAの鎖の交換後結合する一連の反応をCre単独で行っていることが知られている(図1)。最近loxP配列に変異を加えた変異loxP同士では組換えが起きるが、本来のloxP配列とは起こらない変異loxP配列の存在も報告されている。しかしloxP内のスペーサー配列の組換えにおける役割に関しては、まだ十分に解析が行われていない。そこで本研究では、このスペーサー領域の塩基を置換した場合のCreによる組換えへの影響について検討を行った。 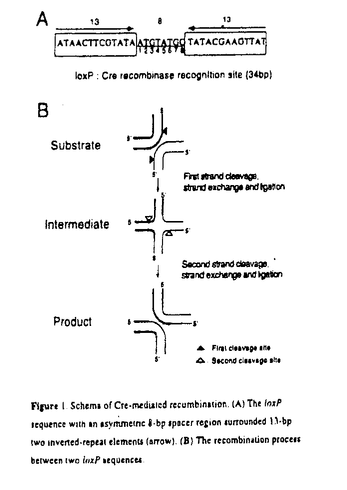 Figure 1 Schems of Cre-meduated recumbination.(A)The loxP sequence with an asymenthc 〓-bp spacer region Surrounded 1)-bp two inverted-repeat elements(arrow).(B)The recombination process between two loxP sequences. Figure 1 Schems of Cre-meduated recumbination.(A)The loxP sequence with an asymenthc 〓-bp spacer region Surrounded 1)-bp two inverted-repeat elements(arrow).(B)The recombination process between two loxP sequences. まず、スペーサー領域の8塩基それぞれを他の塩基で置換した24通りの一塩基変異loxPと2つの塩基を置換した二塩基変異loxPを合成し、in vitro組換え反応系を用いて検討を行った。その時スペーサー領域の左側から塩基に番号(1〜8)を付けた(図1)。従来のin vitro系での変異loxPの研究は変異loxPを合成し、plasmidへ挿入した基質を用いてCreによる組換え反応を行った後、そのsampleを制限酵素で切断し電気泳動することにより反応効率を測定している。本研究では変異loxPを含む合成oligo DNAを、直鎖上に切断したplasmid DNA断片の両側にligationした基質を用いて検討を行った。従来のようにplasmidへ挿入する方法と比較して本法は測定の感度がやや低かったが、大量の変異loxPを一度に解析することができる方法として有用な方法であると考えた。また、一部の実験には従来の方法のようにloxP配列をplasmidへ挿入する方法も併用した。 組換え酵素Creは一般的には大腸菌で発現させ解析に用いている。しかしCreは動物細胞で発現しても大腸菌と同等に機能することが知られており、また組換えアデノウイルスは特に293細胞では大量に目的の蛋白質を生成することが可能な系であるので、Cre発現組換えアデノウイルスを293細胞へ感染し、そのlysateを解析に用いることとした。実際に225cm3のフラスコ1本の感染293細胞から1 gのplasmidをプロセスできる約500回分の活性をもつlysateが得られた。また、そのlysateは論文で報告されている大腸菌由来のものと比較して細胞由来のnucleaseの影響が少なかったため、精製しないでそのままin vitro反応系に用いることが可能であった。 gのplasmidをプロセスできる約500回分の活性をもつlysateが得られた。また、そのlysateは論文で報告されている大腸菌由来のものと比較して細胞由来のnucleaseの影響が少なかったため、精製しないでそのままin vitro反応系に用いることが可能であった。 まずスペーサー領域の塩基を一塩基置換した変異loxPを用いて、本来のloxPと変異loxPの間での組換えと変異loxP同士での組換え反応を行った(図2)。 i)スペーサーの両端の塩基(1番と8番)変異 本来のloxPと変異loxPの間では本来のloxP同士の場合に相等する反応が確認されたが、変異loxP同士は本来のloxPと変異loxPを用いた場合と比較して反応が抑制されていた。そこでスペーサーの両端はCreによる組換え反応において本来のloxP塩基配列そのもの(1:A,8:C)が重要である可能性が示唆された。 ii)スペーサーの左側の塩基(2番から5番まで)変異 変異loxP間では組換えは本来のloxPの場合とほぼ同程度進行することが確認された。また本来のloxPと変異loxPの間では、片鎖のDNAの組換えは起こるが最終産物を生成するためのもう一方のDNA鎖の結合が起こっていない中間体由来のものと思われる生成物が主産物として蓄積し、最終産物はほとんど見られなかった。 iii)スペーサーの右側の塩基(6番、7番)変異 変異loxP間ではii)と同様本来のloxP間の反応と、ほぼ同等の結果が認められた。また本来のloxPと変異loxPの間では中間体も最終産物もわずかな量しか検出されなかった。従って、このsiteは中間体産生よりも前の段階で必要なsiteである可能性が示唆された。 ii)、iii)の結果から、loxPのスペーサー領域に変異があってもCreの認識そのものは本来のloxPと同様に行われること、またスペーサー領域は主にstrand exchangeの段階で重要な役割を果たしていることが示された。更にCreによるstrand exchangeは2段階の反応を経て行われ、最初の一段目の反応である片鎖のstrand exchangeにはスペーサー内の右側の塩基(6,7)が重要であり、二段目の反応には左側の塩基(2,3,4,5)が重要であることも明らかとなった。 そこでこれらの2つの領域に各々1ヶずつ塩基変異を入れた二塩基変異loxPは、本来のloxPとはほとんど組換えを起こさず、変異loxP同士で独立的に組換えを起こす特異性が一塩基変異loxPよりも高いのではないかと考え、検討を加えた。一段目の反応に重要な右側の塩基では、7番の変異の方が6番の変異と比べ本来のloxPとの組換えを抑制していたため、7番の変異と左側の塩基(2,3,4,5)の変異を組み合わせた30通りの二塩基変異loxPを作製した。これらの変異体は7番のみの一塩基変異loxPよりも、同じ変異をもったloxP同士と特異的な組換えが効率よく行われることが確認された。この効率は本来のloxP同士の組換えに匹敵していた。 本来のloxPと変異loxPの間で置換したい遺伝子を挟んだplasmidを細胞内へ導入すると、Creによりdouble reciprocalな組換えによる遺伝子置換が起こることが報告されている。現在この目的で用いられている変異loxPは7番目の塩基を置換した一塩基変異loxP(図2:loxP71)であるが、この二塩基変異loxPを用いることにより、細胞内において、より一層特異性が高く効率の良い遺伝子置換が起こることが期待される。このようなCre/変異loxP系の開発はrecombinaseそのものの研究としてだけでなく、その性質を用いることにより動物細胞での遺伝子置換や、染色体の特定の場所への治療用遺伝子の導入などの遺伝子治療の研究にも応用されるものと考えている。  Figure 2.Effeccts of single-base substitution of the spacer on recombination.Names of loxP mutants and their spacer sequences are shown.The mutated nucleotide is shown in a small letter.The yielded amounts of product and intermediate in molar ratio are representedwhen the amount of wild-type product is regarded as 100%.Similarexperiments were repeated twice to fourtimes and typical results arerepresented.The quantitative results were normally reproduciblewithin±5%.wt,wild-type loxP Figure 2.Effeccts of single-base substitution of the spacer on recombination.Names of loxP mutants and their spacer sequences are shown.The mutated nucleotide is shown in a small letter.The yielded amounts of product and intermediate in molar ratio are representedwhen the amount of wild-type product is regarded as 100%.Similarexperiments were repeated twice to fourtimes and typical results arerepresented.The quantitative results were normally reproduciblewithin±5%.wt,wild-type loxP |