インターフェロン(IFN)は抗ウイルス作用、細胞増殖抑制作用など多彩な生理活性を有するサイトカインである。本研究は、生体内におけるIRFファミリー転写因子p48(ISGF3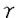 )の詳細な機能を明らかにするために、ジーンターゲティング法を用いてp48欠損マウスを作製し、その解析を試みたものであり、下記の結果を得ている。 )の詳細な機能を明らかにするために、ジーンターゲティング法を用いてp48欠損マウスを作製し、その解析を試みたものであり、下記の結果を得ている。 1. p48遺伝子の翻訳開始コドンを有するエクソンおよび第2エクソンをネオマイシン耐性遺伝子に置換したターゲティングベクターを構築し標的組換えを試みた結果、140個のG418耐性クローンより相同組換えの起こった2クローンを選抜した。これらの相同組換え体からp48遺伝子欠損マウスを作製した。p48遺伝子欠損マウスは同腹野生型マウスと比較して、大きさ、体重、行動および生殖能においてその差異は認められなかった。 2. p48欠損マウス由来の胎児線維芽細胞(EFs)を用いて、3種類のウイルス,EMCV,VSVとHSVに対する抵抗性をプラークアッセイ法によって調べた。p48欠損EFsではIFN-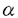 のみならずIFN-によるEMCVの抗ウイルス作用の誘導がほぼ完全に障害された。また、VSVおよびHSVに対する抗ウイルス作用についても、p48欠損EFsは野生型と比べウイルス増殖抑制効果の顕著な低下が認められた。これらの結果より、p48がI型IFN応答のみならずII型IFN応答においても必須であることが強く示唆された。 のみならずIFN-によるEMCVの抗ウイルス作用の誘導がほぼ完全に障害された。また、VSVおよびHSVに対する抗ウイルス作用についても、p48欠損EFsは野生型と比べウイルス増殖抑制効果の顕著な低下が認められた。これらの結果より、p48がI型IFN応答のみならずII型IFN応答においても必須であることが強く示唆された。 3. 抗ウイルス状態の確立に重要と考えられる種々のIFN誘導遺伝子のp48に対する依存性を調べた結果、IFN-およびIFN-による2-5oligo adenylate synthetase(2-5A合成酵素)およびdouble-stranded RNA-dependent protein kinase(PKR)のIFN誘導遺伝子群のmRNA発現は、完全にp48に依存性であった。 4. p48がIFN-によって誘導されるIFN誘導遺伝子の転写活性化にどのように関わっているかを調べた結果、IFN-のみならずIFN-によってもISGF3そのものが直接的に誘導されることが示された。さらに、ISGF3の構成要素であるStat2がIFN-同様、IFN-によってもリン酸化されることを証明した。 5. IRF-1とp48,両因子がどのように相関してIFN作用を調節しているかを詳細に検討するためにIRF-1,p48両遺伝子欠失マウス(dko)を作製した。その結果、IFN-によるMHCクラスI,TAP1,LMP2遺伝子の発現は、IRF-1欠損ないしp48欠損細胞では野生型に比べ減少していたが、dko細胞では殆ど完全に消失していた。さらに、dko細胞では細胞表層のIFNによるクラスI分子発現の増強が完全に喪失しており、無処理のものと同等であった。従って、MHCクラスI依存性抗原提示機構は、II型IFN作用によって増強される過程においてIRFファミリー転写因子であるIRF-1とp48(ISGF3)の相補的な作用が重要であるとが示唆された。 以上、本論文はIRFファミリー転写因子p48がI型およびII型IFNによる抗ウイルス状態の確立に必須であること、そしてMHCクラスI遺伝子をはじめとする抗原提示機構に重要と考えられるIFN誘導遺伝子群の発現がIRF-1とp48の相補的な作用に依存していることを明らかにした。本研究は、転写活性化因子による遺伝子発現制御という見地に基づいたアプローチによりウイルス感染に対する生体防御機構の解明に重要な貢献をなすと考えられ、学位の授与に値するものと考えられる。 | 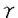 )の役割
)の役割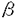 )はウイルスやpolyd(I):d(C)など二重鎖RNAによってさまざまな細胞で産生される。一方、II型IFN(IFN-
)はウイルスやpolyd(I):d(C)など二重鎖RNAによってさまざまな細胞で産生される。一方、II型IFN(IFN-