血管内皮細胞(ECs)から産生されるnitric oxide(NO)は血管拡張作用を有し、平滑筋細胞(VSMCs)の増殖を抑制する。NOの産生異常は高血圧及び動脈硬化の発症に重要な役割を果たす。また、NOの産生はECs内のCa2+シグナルによって制御される。G蛋白質を細胞内信号伝達に用いる場合、典型的なCa2+の経時変化は各種の刺激によりまず細胞内Ca2+(Ca2+i)濃度が急速に上昇し、peakに到した後、徐々に下降する。最初のpeakはIP3-induced Ca2+ release(IICR)を介する細胞内ストアからのCa2+放出で、下降相は細胞外からのcapacitative Ca2+ entry(CCE)で構成される。しかし、両者のどちらがNO産生に重要なのか不明である。NOの正確な定量が難しく、従来NO-donorの作用は不確定であるため、NOの産生量とVSMCs内のCa2+動態の関係も不明であった。一方、産生されたNOはautocrine作用でECs自身のCa2+iを下げ、IICRに影響する。ECsの細胞膜上の数種の受容体に対するagonist、G蛋白質及PLCなどの特異的なantagonistが少なく、また各々の細胞に選択的に使用できないため細胞間の比較ができない。NOのautocrine作用を正確に評価する事が難しかった。NOがECs自身のCCEに及ぼす影響について全く不明である。従って、本研究の目的は:1)NO産生時のECs内Ca2+シグナルとNOの産生量、共存培養してECsに隣接するVSMCs内Ca2+動態に及ぼす影響を解明する。2)ECs内IICRのNO産生過程における意義を解明する。3)IICRを抑制してNOのautocrine作用を正確に定量する。4)産生されたNOがIICRとCCEに及ぼす影響を検討する。以下の結果を得ている。 1.NO産生の経時変化、Ca2+流入量によるNO産生量の変化: 牛大動脈から分離したECsを単独培養し、4種の異なった細胞内信号伝達系を有する刺激物質、即ちATP(10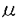 M)、bradykinin(BK,100nM)、ionomycin(IM,50nM)またはthapsigargin(TG,1M)をECsに加え、刺激前後Ca2+i濃度変化を二次元画像法で解析した。同時に刺激後培地中に放出されるNO2の代謝物質NO2をchemiluminescence法で測定した。ATPとBKは刺激後3分間でNO産生が安定化し、IMは20分後に定常化した。TGは40分まで増加した。上記4種の刺激物質とも、NO産生量は細胞外に生理的な濃度のCa2+が存在した時増加し、その量はTG>IM>ATP>BKの順番に減少した。細胞外のCa2+をEGTA-でchelate又は細胞内へのCa2+流入をNi2+で遮断するとNO産生量は著明に減少した。 M)、bradykinin(BK,100nM)、ionomycin(IM,50nM)またはthapsigargin(TG,1M)をECsに加え、刺激前後Ca2+i濃度変化を二次元画像法で解析した。同時に刺激後培地中に放出されるNO2の代謝物質NO2をchemiluminescence法で測定した。ATPとBKは刺激後3分間でNO産生が安定化し、IMは20分後に定常化した。TGは40分まで増加した。上記4種の刺激物質とも、NO産生量は細胞外に生理的な濃度のCa2+が存在した時増加し、その量はTG>IM>ATP>BKの順番に減少した。細胞外のCa2+をEGTA-でchelate又は細胞内へのCa2+流入をNi2+で遮断するとNO産生量は著明に減少した。 2.単独培養のECsのCa2+i動態とNO産生量の関係: ECs内Ca2+動態を定量する為、peakの濃度、peak後の下降相の半減期およびCa2+i動態の時間積分を測定した。4種の刺激物質とも細胞外にCa2+が存在した時にCa2+i濃度は増加し、peakの値はBK>ATP>IM>TGの順番に減少した。細胞外Ca2+をEGTAでchelate、又はCa2+流入をNi2+で遮断した結果、4種の刺激物質ともCa2+iのpeakレベルは不変で、下降相の半減期及びCa2+i濃度の時間積分値が減少した。NO産生量はCa2+i上昇のpeak後の下降相の半減期(r=0.90)及びCa2+i濃度時間積分(r=0.86)とは良い指数関係を示した(p<0.001)。 3.単独および共存培養のVSMCsのCa2+i反応: 単独培養のVSMCsでは、ATP、TGやIMはCa2+iを増加させたが、BKによる増加分は少なかった。ECsとVSMCsを共存培養し、そのECsから産生されたNOによって隣接するVSMCs内Ca2+濃度の動態を定量観測した。共存培養では、4種の刺激物質ともVSMCsのCa2+i濃度を逆に低下させた。その低下程度とECsからのNO産生量は良く相関し、TG>IM>ATP>BKの順であった。Indomethacin又はgly benclamide前処理では全くVSMCsのCa2+i動態に影響が認められなかったが、l-NMMAの前処理でCa2+i濃度の低下は阻止された。これは共存培養のVSMCsのCa2+i反応がNOにより修飾された事を示す。 4.L-arginineの結果: ECsの単独培養で、特に刺激物質を加えなくとも高濃度のl-arginine(5mM)はNO産生を促進した。且つこの促進作用は細胞外のCa2+有無に影響されなかった。共存培養でl-arginineはECs内Ca2+濃度を軽度に低下させ、VSMCsのCa2+i濃度を大幅に低下させた。 5.ECsのIP3受容体subtypeの検出とMicroinjectionの対照実験: 上記の結果で、CCEはNO産生に重要であることを証明した。しかし、その時のECs内IICRのNO産生過程における生理的意味は不明である。従って、ヒトの1(IP3R1)、2(IP3R2)、3(IP3R3)型IP3受容体のC末にあるCa2+ channeldomainに対する特異的な抗体(IP3R1Ab,IP3R2Ab,IP3R3Ab)を作りaffinity chromatography法で精製し、immunoblotting法で特異性を確認した。ECsではIP3R1しか存在しないことを示した。又、ECs-をryanodineで前処理した後、caffeineに対して弱いCa2+i反応をしか示さず、Ca2+-induced Ca2+ release-の機構は重要でないことを確認した。Normal IgGと抗体のvehicleをmicroinjectionした細胞はATPに対するCa2+i反応が全く影響されなかった。30分後に二回目のATP刺激を行い、初回と同様のCa2+i反応が示された。 6.IP3R1Ab又はheparinによるIICRの抑制: IP3R1Abを個々のECsにmicroinjectionした後、二次元画像法でATPの刺激に対するIICRとCCE両者の反応が抑制された。Microinjectionされた細胞はIM又はTGに対して正常な反応を示し、microinjectionによる細胞傷害は起こさないことを示唆した。小量のheparin(MW=5,0000)で軽度IICRを抑制し、Ca2+iの上昇は遅くなり、CCEの段階のCa2+i濃度は低下した。多量のheparinでIICRを完全に抑制し、更にCa2+iを基線以下-51%位まで低下した。L-NMMA前処理でCa2+i濃度を低下させず、基線以下まで低下したCa2+i濃度はATP刺激後、周囲の細胞から放出されたNOによることを示唆した。 7.NOによるIICRとCCEの修飾およびCCE過程中のNO産生: 抗体等の操作を加えていない細胞をATP、BK又はTGで刺激する過程中のCa2+i濃度上昇とMn2+ quenchingにおける内因性NOと外因性NO donorの修飾作用を記録した。L-NMMAで前処理すると、ATP又はBKの刺激で産生した内因性NOがIICRを20%程度まで抑制した。内因性NOを抑制した後、更に外因性NO donorを加え、内因性NOの機能を再現することができた。非常に興味有る事に、NOがCCEを上昇させる作用をもち、その作用はMn2+ quenchingの記録からも証明した。L-NMMAとG-kinase遮断薬、LY83583、はCCE-を抑制させ、逆にNOR-3とBr-cGMPはCCEを亢進させたことから、NOのCCEを促進する機構がG-kinaseを介することを示唆した。TGでCCEを促進すると、大量のNOが産生された。そのNOの産生がl-NMMAの前処理で完全に抑制された。 以上、ECsからのNO産生量が持続性Ca2+流入で決められ、そのNOは隣のVSMCsのCa2+iの下げる程度を濃度依存的に制御する。高濃度の基質があればECsのCa2+iが増加しなくてもNOは産生される。IP3R1を介するIICRはCCEの誘発とその後のNO産生の重要なstepである一方、NOの強いautocrine作用を部分的に緩衝する。NOとCCEの間にはpositive feedbackの機構が存在する。この事からCCEによる大量のNO産生が説明される。NOはIICRを抑制し、CCEを促進することでECsのCa2+i濃度の急速な変化を緩衝することが示された。本論文はECsにおけるNOの産生機構とCa2+i動態の関係解明に重要な貢献をなすと考えられ、学位の授与に値するものと考えられる。 |