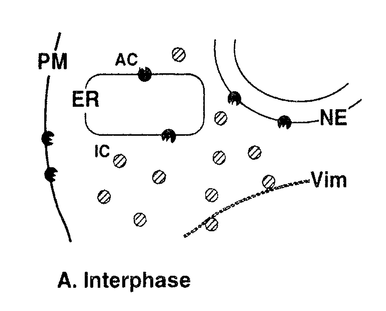
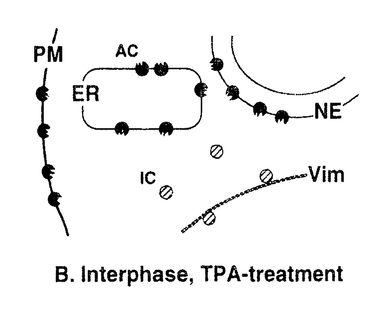
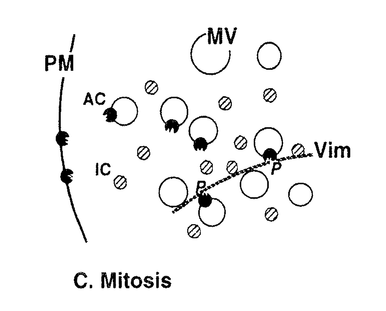
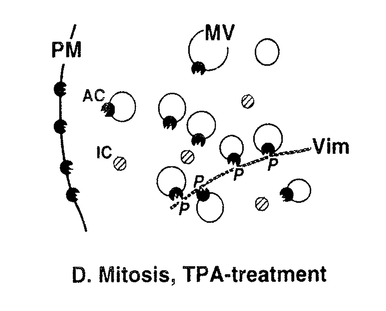
本研究は、細胞周期におけるCキナーゼによる細胞骨格タンパク質・ビメンチンのリン酸化の有無とその制御メカニズムを明らかにするために、Cキナーゼによる特異的なリン酸化部位の1つである、ビメンチン-セリン33(Vim-Ser33)のリン酸化状態を特異的に認識するモノクローナル抗体群を作製し、これを用いて細胞分裂期のビメンチンリン酸化について解析したものであり、下記の結果を得ている。 1.モノクローナル抗体YT33は、Cキナーゼによりリン酸化されたビメンチンを特異的に認識する。本抗体を用いてヒトグリオーマ細胞を免疫染色すると、間期細胞ならびに分裂前期・後期の細胞は染色されず、分裂中期および後期の細胞が特異的に染色された。この染色像はcdc2キナーゼによるビメンチンリン酸化像とは異なり、別の分裂期ビメンチンキナーゼ(おそらくCキナーゼ)が分裂中期特異的にビメンチン分子上のSer33残基をリン酸化することを示唆した。また、同じくCキナーゼによるリン酸化部位であるビメンチン-セリン50(Vim-Ser50)のリン酸化状態を特異的に認識する抗体TM50を用いても、同様の染色像が観察された。更に、Cキナーゼの活性化剤として知られるphorbol ester(TPA)で細胞を刺激したところ、分裂期細胞ではVim-Ser33のリン酸化量の増大が観察された。一方、間期細胞ではTPA刺激後もVim-Ser33のリン酸化は観察されず、Cキナーゼによるビメンチンリン酸化は、in vivoでは分裂期細胞に特異的な現象であることが示唆された。抗体TM50を用いたTPA刺激実験でも同様の現象が観察された。 2.従来より、Cキナーゼは細胞内で活性化されると細胞質から細胞膜への移行(translocation)を起こし、そこで細胞膜を構成するリン脂質依存性にリン酸化活性を示すことが知られている。間期細胞および分裂期細胞を細胞質画分と細胞膜画分に分け、それぞれに含まれるCキナーゼ活性を測定すると、両者の細胞内局在の差異は検出されず、分裂期特異的なCキナーゼの活性化は認められなかった。 3.Cキナーゼによるビメンチンリン酸化はin vivoにおいて分裂期特異的な現象であったが、細胞を超音波処理により破砕して調製したwhole cell lysateを用いたin vitroのリン酸化実験では、間期細胞、分裂期細胞ともにVim-Ser33リン酸化活性が認められた。更にTPA処理した細胞では、間期・分裂期ともにVim-Ser33リン酸化活性の増大が認められた。 以上、本論文は新たに作製したモノクローナル抗体群を用いて分裂期細胞における細胞内シグナル伝達を解析し、Cキナーゼが分裂期特異的なビメンチンキナーゼとしてフィラメント構築の制御に関与すること、また、このビメンチンリン酸化には、細胞分裂に伴う細胞内膜構築の再編成が新たな制御メカニズムとして関与することを世界に先駆けて発見した。本研究は細胞分裂期におけるCキナーゼの機能とその制御機構の解明に重要な貢献をなすと考えられ、学位の授与に値するものと考えられる。 |